|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
КЛАСС I. МНОГОНОЖКИ (MYRIAPODA)
Класс Myriapoda включает около 10000 видов исключительно наземных, иногда довольно крупных членистоногих. Червеобразное, вытянутое тело многоножек подразделяется на два отдела: слитную голову и чле-
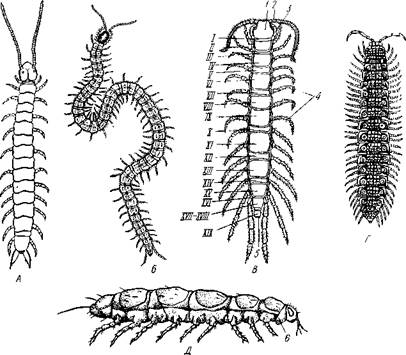
Рис. 301. Различные многоножки (из Беклемишева): А — Scolopendrella immaculata (подкл. Symphyla); Б — Pachimerium ferrugineum (подкл. Chilopoda); В — Lithobius forficatus (подкл. Chilopoda); Г — Polydesmus complanatus (подкл. Diplopoda); Д — Pauropus silvaticus (подкл. Pauropoda): 1 — голова, 2 — антенна, 3 — ногочслюсть, 4 — туловище, 5 — анальное отверстие, I—XIX туловищные сегменты, 6 — шейный сегмент, гомологичный сегменту мак- снлл II нистое туловище, которое часто состоит из значительного числа снабженных конечностями сегментов. Все многоножки объединяются в 4 подкласса: симфилы (Symphyla), пауроподы (Pauropoda), двупарноногие, или кивсяки (Diplopoda), и гу-боногие (Chilopoda). Строение и физиология.Тело многоножек (рис. 301) состоит из резко
отделенных друг от друга сегментов, число которых варьирует в широких пределах: от 18 у симфил (подкл. Symphyla) и 14 у пауропод (подкл. Pauropoda) до 181 у некоторых губоногих (подкл. Chilopoda). Голова Myriapoda ясно обособлена от туловища. Она включает акрон и слившиеся с ним 4 (подкл. Symphyla и Chilopoda) или 3 (подкл. Pauropoda и Diplopoda) первых сегмента тела. Во втором случае последний головной сегмент остается свободным и называ-
Рис. 302. Ротовой аппарат Lithobius for-ficatus (Chilopoda) (из разных авторов). А — внешний вид головы; Б — мандибула— верхняя челюсть; В — максиллы I; Г — максиллы II: 1 — основание усика. 2 — мандибула, 3 — максилла I, 4 — основание туловищной ножки, 5 — ногочелюсть, 6' — максилла II Рис. 303. Ротовой аппарат Diplopoda (из разных авторов). А — внешний вид головы; 5 — мандибула; В — гнатохилярий: / — основание усика, 2 — мандибула, 3 — гнатохилярий
ется «-шейным». Эта особенность строения головы части многоножек справедливо рассматривается как примитивный признак. Голова несет усики и ротовые конечности: верхние челюсти — мандибулы, или жвалы, и нижние челюсти — максиллы, которых соответственно числу сегментов, вошедших в состав головы, может быть одна или две пары. Усики, или антенны, многоножек отвечают антеннулам (антеннам I) раков и относятся к акрону. Они более или менее длинные, тонкие, разбиты на членики и большей частью неветвистые. По-видимому, они служат органами не только осязания, но и обоняния. Конечности, соответ ствующие антеннам II раков и принадлежащие первому сегменту тела4 у многоножек редуцированы. Но их сегмент, называемый пнтеркаляр-ным, имеется. Прочие головные конечности превращены в ротовые органы и гомологичны соответствующим ротовым частям раков. Строение их неодинаково в разных подклассах многоножек. У представителей подклассов Symphyla и Chilopoda (рис. 302) рот спереди прикрыт хити-низированной складкой покровов — верхней губой; последняя по происхождению не имеет ничего общего с конечностями. Мандибулы, конечности второго сегмента, состоят из двух коротких жевательных пластинок с зазубренным внутренним краем. Первая и вторая пары нижних челюстей (конечности III и IV сегментов соответственно) у большинства представителей состоят из основания, на котором сидят членистый челюстной щупик и нерасчлепенные жевательные лопасти. И щупик и лопасти могут частично редуцироваться. У представителей подклассов Pauropoda и Diplopoda (рис. 303) за верхней губой и парой мощных зазубренных мандибул располагается всего одна непарная пластинка — гнатохилярий (gnathochilarium) довольно сложного строения. История развития показывает, что она закладывается в виде парного зачатка и соответствует первой паре нижних челюстей губоногих. Конечности шейного сегмента редуцировались1. За головой следует большей частью однородно построенное туловище. Однако строго гомономная сегментация бывает выражена только у самых примитивных форм (см. рис. 301,Б). В процессе эволюции характер сегментации заметно меняется. У некоторых многоножек (ряд губоногих) часть туловищных сегментов заметно уменьшается в размерах (см. рис. 301,6). При этом уменьшенные и нормальные сегменты довольно правильно чередуются. Иная картина наблюдается у представителей подкласса Diplopoda, у которых происходит попарное слияние большей части сегментов (кроме первых четырех, включая и «шейный» сегмент). Каждый такой двойной сегмент соответственно несет не одну, а две пары конечностей (рис. 304). Подобные отступления от исходной примитивной гомономности не ведут, однако, к подразделению туловища на тагмы. Лишь у кивсяков первые туловищные сегменты, несущие по одной паре конечностей (рис. 304) 'и тем отличающиеся от прочих, вместе с безногим «шейным» сегментом иногда обозначаются как «грудные», а следующие за ними двойные сегменты — «брюшные». Однородность туловищных сегментов многоножек определяет и сходство строения их конечностей, которые имеют вид простых ходных ножек, состоящих из одного ряда члеников и заканчивающихся коготком. Примеры их функциональной и морфологической дифференциации не- 1 Существует и другая точка зрения, согласно которой гнатохилярий Diplopoda образуется за счет максиллИ. Шейный сегмент в этом случае рассматривается как первый туловищный сегмент. многочисленны. Так, у губоногих характерно превращение ножек первого туловищного сегмента в ногочелюсти (см. рис. 302,Л), играющие главную роль в захвате и убивании добычи. Эта пара ножек сильно увеличивается в размерах и обладает крайне утолщенным основным члеником, тогда как концевой членик сильно заострен и загнут в виде крюч-
Рис. 304. Передний и задний концы тела с левой стороны самца кивсяка Schizophyllum sabulosum (по Иванову): / — голова, 2—глазное поле, 3—антенна, 4—обонятельные органы, 5 —верхняя губа, 6 — верхняя челюеть, 7 — щека, 8 — видоизмененная ножка 1-го туловищного сегмента, сместившаяся вперед. 9 — ножка 2-го туловищного сегмента, сместившаяся вперед, 10 — место полового отверстия. И — ножка 3-го сегмента, 12 — ножки двойных сегментов, 13 — тельсон, 14 — «молодые» сегменты зоны роста, /Л — сегменты, слившиеся в двойной сегмент, 16 — отверстия ядовитых желез, 17—«шейный» сегмент, соответствующий сегменту максплл И; I — 1П — сегменты «груди»; IV. V, VI н т. д. — сегменты «брюшка» ка. В основании конечности залегает ядовитая железа, проток которой открывается на конце крючка. Выделяемый яд сильно действует на членистоногих и позвоночных. Укус пальца крупной сколопендрой (Scolo-pendra) приводит к временному опуханию всей руки. Некоторые пары ножек, принимающие участие в копуляции, несколько изменяются в строении и называются гоноподиями. Тело покрыто хитиновой, иногда пропитанной известью кутикулой, выделяемой однослойным гиподермальным эпителием. Последний довольно богат одноклеточными и многоклеточными кожными железами, среди которых особенно интересны защитные железы кивсяков. Они помещаются на спинной стороне части туловищных сегментов и открываются наружу оборонительными отверстиями, из которых выпрыскивается секрет. У разных видов подкласса Diplopoda секрет необычайно варьирует по виду и химическим свойствам. Так, секрет Spirobolus едок и окрашивает кожу человека в темный цвет. Polyzonium rosalbum выделяет млечную жидкость, имеющую запах и жгучий вкус камфоры. Тропическая Fontarla содержит в железах свободную синильную кислоту и пахнет горьким миндалем. Пищеварительная система многоножек (рис. 305) имеет вид прямой трубки; лишь в области задней кишки пищеварительный канал образует петлевидный изгиб. Рот лежит на брюшной стороне головы между ротовыми конечностями и ведет в переднюю кишку, часто называемую пищеводом. С начальным отделом пищеварительной системы связаны слюнные железы. Кив-сяки обладают тремя парами желез, открывающимися самостоятельными протоками в ротовую полость и у основания гнатохиляриума. Ввиду образования их из мезодермы эти железы считают видоизмененными целомодуктами. У губоногих 3—5 пар слюнных желез с самостоятель- ными протоками, открывающимися в ротовую полость или по бокам рта. Они, по-видимому, чисто эктодермального происхождения, т. е. представляют модифицированные кожные железы. Слюнные железы, открывающиеся на второй паре челюстей, приравнивают к прядильным желе-
Рис. 305. Внутренние органы многоножек. А—пищеварительная система Lithobius fofficalus (Chilopoda) (по Плате); Б — мужская половая система Chilopoda (из Шауфлера); В — женская половая система Chilopoda (из Шауфлера); Г—мужская половая система Diplopoda (по Фабру); Д — женская половая система Diplopoda (по Фабру): 1 — пищевод, 1 — средняя кишка, 3 — задняя кишка, 4—анус, 5 — мальпигиевы сосуды, в — слюнная железа, 7 — семенник, 8 — семенной пузырек, 9 — парный участок семяпровода, 10 — семяизвергательный канал, // — придаточные железы, 12 — яичник, 13 — непарный участок яйцевода, 14 — парный участок яйцевода, 15 — половое отверстие, 16 — половая ножка, 17 — семяприемник, 18 — придаточные железы. 19 — семяпровод, 20 — вскрытый половой мешок зам личинок насекомых, отверстия которых помещаются на той же паре ротовых конечностей. Средняя кишка служит местом переваривания и всасывания пищи. Задняя кишка короткая. Представители подкласса Diplopoda растительноядны и питаются главным образом гниющими листьями, растительными остатками, древесной трухой и т. п. Губоногие — хищники, питающиеся насекомыми. Выделительная система. На границе между средней и задней кишками в кишечник впадают 1 или 2 пары (последнее у Chilopoda) длинных слепозамкнутых на свободном конце трубок — мальпигиевых сосудов (рис. 305,Л). В эпителии сосудов и их просвете скапливаются конкреции мочевой кислоты; последняя у многоножек, так же как и у насекомых (с. 352), представляет собой главный продукт выделения. Кроме мальпигиевых сосудов выделительную роль играют и другие образования, прежде всего лимфатические железы в виде неправильных клеточных тяжей, расположенных либо вдоль мальпигиевых сосудов, либо вдоль брюшного кровеносного сосуда, или вдоль брюшной нервной цепочки. Они служат для захвата и накопления твердых продуктов выделения и фагоцитнруют впрыснутые в полость тела твердые частицы (например, порошка туши или кармина). Кроме того, участие в выделении прини- мает жировое тело. Полость тела — миксоцель — многоножек во многих местах заполнена неправильными тяжами и скоплениями клеток, причем эти скопления ограничены тонкой собственной оболочкой. Совокупность таких скоплений и называется жировым телом. В клетках его наблюдаются многочисленные капли жира, а также конкреции мочевой кислоты. Жировое тело служит не только для накопления запасного питательного материала, но и для выделения (мочевая кислота). Нервная система состоит из головного мозга, окологлоточных кон-нективов и брюшной нервной цепочки. Головной мозг имеет довольно сложное гистологическое строение, свидетельствующее, что голова многоножек образована из сегментов, превышающих по числу количество пар головных придатков. В мозге помимо пары ганглиев, посылающих нервы к антеннам, имеются еще парные скопления нервных клеток, отвечающие вставочному (интеркалярному) сегменту. Брюшная цепочка состоит из расположенного в голове подглоточного ганглия, который инпервирует все ротовые конечности, и из длинного ряда туловищных ганглиев, хорошо обособленных и сидящих на общем парном продольном нервном стволе. Па каждый сегмент приходится обычно один парный ганглий. У Diplopoda такое устройство наблюдается лишь в четырех передних сегментах, прочие же содержат по два расположенных один за другим ганглия, чем доказывается сложный состав этих сегментов. Органами осязания и обоняния служат антенны, усаженные чувствительными волосками, чувствительными колбочками и т. п. Кроме того, по бокам головы, между основаниями антенн и глазами, лежат два темсшваровых органа чувств (по-видимому, хеморецепторы). Это или подковообразные ямки, на дне которых имеются валики из чувствительных клеток, или же скопления чувствительных клеток, помещающиеся под покровами головы в глубине длинных узких каналов. Темешваровы органы иннервируются от головного мозга. Громадное большинство многоножек снабжено глазами, их может быть 2, 4 или много. Глаза сидят по бокам головы и носят характер одиночных, просто устроенных глазков. Только у мухоловок (Scutigera) на голове два больших скопления глазков, так тесно сближенных, что они соприкасаются друг с другом и напоминают сложные фасетированные глаза насекомых. Зрительные способности многоножек невелики. Многоножки предпочитают затененные участки. Органы дыхания представлены трахеями — тонкими воздухоносными трубочками эктодермального происхождения, возникающими как глубокие впячивания покровов. Стенки трахей выстланы продолжением наружной кутикулы, которое на всем протяжении трахейной трубки образует спиральное утолщение, не дающее трахеям спадаться. Трахеи начинаются парными дыхальцами, или стигмами, лежащими на брюшной стороне сегментов туловища. Как исходная форма трахейной системы должна рассматриваться такая, у которой каждый туловищный сегмент несет пару стигм, а каждая стигма ведет в обособленный пучок тонких трахейных трубочек. Ближе всего к этой схеме стоит подкласс Diplopoda, у представителей которого почти все туловищные сегменты снабжены независимыми друг от друга парными пучками неразветвленных трахей. В связи с двойным характером туловищных сегментов последние несут не по 1, а по 2 пары дыхалец. У большинства видов подкласса Chilopoda стигмы расположены на туловище через сегмент, а у некоторых форм (например, Scutigera) животное имеет всего 7 пар стигм, но сама трахейная сеть развита у губоногих гораздо сложнее. Трахеи у некоторых из них сильно ветвятся, и между трахейными пучками соседних сегмен- тов и одного и того же сегмента (правым и левым) устанавливаются сообщения в виде продольных и поперечных перемычек. Концевые веточки трахеи многоножек оплетают все внутренние органы. Смена воздуха в трахеях происходит вследствие изменения объема тела при сокращении и расслаблении мускулатуры. Кровеносная система довольно хорошо развита, кроме сердца есть система периферических кровеносных сосудов. Сердце в виде нежной прозрачной трубки тянется над кишкой вдоль всего туловища и сзади замыкается слепо или же продолжается в два коротких сосуда, теряющихся в мускулатуре. Сердце соответственно сегментам поделено на камеры: каждая камера имеет две остии. У подкласса Diplopoda, где сегменты двойные, остий по две пары на один сегмент. Сердце продолжается в головную аорту, направляющуюся к мозгу. У губоногих кровеносная система сложно устроена: аорта отдает от себя на пути к мозгу артериальное кольцо, огибающее кишку и впадающее в брюшной продольный сосуд, лежащий над брюшной нервной цепочкой. От каждой камеры сердца отходят, кроме того, 2 боковые артерии. Сердце подвешено к стенкам тела при помощи особых крыловидных мышц. Вес отходящие от сердца сосуды более или менее богато ветвятся, но затем обрываются, и гемолимфа попадает в лакуны миксоцеля, т. е. в промежутки между органами. Из лакун она поступает в околосердечный участок полости тела и оттуда вновь в сердце. Сердце гонит гемолимфу от заднего конца к переднему, в брюшном сосуде она движется в противоположном направлении. Половая система. Все многоножки раздельнополы. Половые железы лишь в редких случаях сохраняют (некоторые пауроподы) первоначально парный характер и обычно сливаются в непарное образование различного вида. Так, семенник, например, имеет вид массивного образования с лопастями по краю или длинной тонкой трубки, или состоит из 11—12 пар маленьких долек, соединенных общим половым протоком. Яйцевод и семяпровод у кивсяков (см. рис. 305, Г, Д) в начальной части представляют непарные протоки. Направляясь вперед, они раздваиваются и открываются наружу на брюшной стороне второго (не считая шейного) туловищного сегмента. На этом же сегменте расположено половое отверстие у симфил и пауропод. У представителей подкласса Chilopoda половой проток в начальной части непарный, может образовывать две ветви, которые затем обязательно сливаются. Половое отверстие расположено на предпоследнем сегменте .туловища (см. рис. 305, Б, В). С половой системой многоножек связан ряд дополнительных образований. Так, в семяпровод часто впадают длинные мешковидные семенные пузырьки. Женская половая система может быть снабжена семяприемниками. Часто развиваются особые придаточные железы (см. рис. 305, Б, В). Способы оплодотворения многоножек разнообразны. В более простом случае самец подвешивает на выделенную им паутину капельку семенной жидкости или настоящий сперматофор, которые позднее подбираются самкой. Иногда происходит копуляция, причем семенная жидкость в таком случае вносится в половое отверстие самки конечностями самца (чаще всего для этого служат специализированные конечности — гоно-подии). Развитие.Яйца многоножек крупны и богаты желтком, соответственно с чем они испытывают частичное, поверхностное дробление. Постэмбриональное развитие Myriapoda может протекать по двум несколько различным типам, Первый тип, или настоящее прямое развитие, встречается у части представителей подкласса Chilopoda (Geophilus, Scolopendra): молодое животное вылупляется из яйца, обладая полным числом туловищных сегментов и конечностей, т. е. вполне походит на материнский организм. Второй тип, или развитие с анаморфозом, имеется у прочих губоногих и
двупарноногих. В этом случае животное вылупляется с неполным числом туловищных члеников, восполняемых при ряде линек. С каждой линькой к имеющимся сегментам прибавляются позади последнего сформированного сегмента членики, следующие за ним по порядку. Образование их идет за счет зоны роста, которая лежит непосредственно впереди тельсона (т. е. там же, где и у личинок раков). Молодь анаморфных видов подкл. Chilopoda вылупляется с 12 парами туловищных конечностей, молодь подкл. Diplopoda — всего с 3 передними парами ходных ножек, за которыми следует несколько безногих сегментов. Эта шести-ногая стадия (рис. 306) напоминает личинок многих насекомых, когда они еще лишены зачатков крыльев. Экология.Многоножки преимущественно ночные, избегающие дневного света животные, прячущиеся под корой, камнями и т. п. Кивсяки очень неповоротливы и медлительны, тогда как губоногие, наоборот, ловки и отличаются быстротой движений. Многие многоножки обнаруживают заботу о потомстве. Они или откладывают яйца в особые гнездышки, приготовленные из земли или другого материала, или свертываются спирально вокруг отложенной кучки яиц и остаются в таком положении по нескольку недель, не принимая пищи, пока не вылупится молодь. На севере разнообразие многоножек невелико. На юге — в Крыму, на Кавказе, в Средней Азии количество видов их увеличивается. Самые крупные сколопендры и кивсяки — до 28 см длиной и в палец толщиной— встречаются лишь под тропиками. Наиболее мелкие многоножки имеют всего 1—3 мм в длину. Все многоножки, кроме подкл. Chilopoda, совершенно безобидны. Укусы крупных губоногих, например Scolopendra, могут быть болезненными. Классификация.Многоножки делятся на 4 подкласса (иногда им придают значение самостоятельных классов): симфилы (Symphyla), пауро-поды (Pauropoda), двупарноногие, или кивсяки (Diplopoda), и губоногие (Chilopoda).
Поиск по сайту: |