|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Этапы пути инициации репликации на ОНР у дрожжей
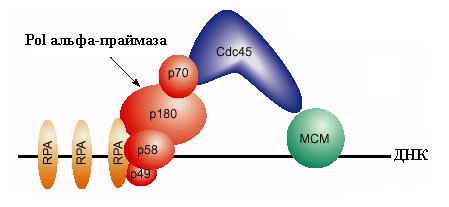
В конце митоза или в начале фазы G1 клеточного цикла нуклеопротеиновые комплексы ORC-ARS вербуют на ДНК белок Cdc6 c мол. массой 58 кД. Этот белок очень нестабилен и должен синтезироваться de novo после выхода клеток из митоза. Он имеет несколько доменов гомологии с субъединицами Orc1, Orc4 и Orc5 комплекса ORC и взаимодействует с ним. Кроме того, Cdc6 гомологичен белкам g-комплекса бактериальной ДНК-полимеразы III и эукариотического комплекса RFC – многосубъединичным ферментам, катализирующим зависящую от гидролиза АТФ погрузку на ДНК скользящих зажимов ДНК-полимераз. Подобно этим ферментам класса АТФаз ААА+, белок Cdc6 способен связывать и медленно гидролизовать АТФ in vitro. Консервативный домен связывания нуклеотидов в Cdc6 необходим для функционального взаимодействия с ORC-ARS in vivo. По аналогии с погрузчиками факторов процессивности, считается, что Cdc6 является погрузчиком на хроматин кольцевого комплекса MCM – ДНК-геликазы репликативных вилок, концентрация которой в клетках дрожжей в 10-100 раз больше концентрации ORC. Белок Cdc6 преимущественно взаимодействует в ORC с комплексом Orc1-АТФ. После узнавания ORC белком Cdc6, связавшим АТФ, образуется комплекс ARS-ORC-Cdc6, который, используя катализируемый Cdc6 гидролиз АТФ, привлекает к ОНР белки Mcm2-Mcm7. В этом процессе у S. cerevisiae участвует также не гомологичный субъединицам гексамера МСМ белок Mcm10, взаимодействующий с Mcm7. Отметим также, что связанный с ORC белок Cdc6 cпособствует ассоциации фактора транскрипции Abf1 c элементом В3 в ARS. Погрузка геликазы МСМ на ORC-ARS в начале фазы G1 завершает образование “предрепликативного комплекса”, в котором ОНР получила “лицензию на репликацию” и перешла в компенентное для инициации состояние. Однако “запуск” (firing) репликации на уже готовой к инициации ОНР откладывается до фазы S клеточного цикла. Для такого запуска необходимо действие циклин-зависимых протеинкиназ, которые появляются только в начале этой фазы. К ним относятся комплексы главной киназы Cdc28 c циклинами типа В (Clb5 и Clb6) и киназы Cdc7 c её регуляторным циклиноподобным белком Dbf4. Циклины Clb5 и Clb6 синтезируются уже в фазе G1 и ассоциируются с Cdc28, но эти комплексы остаются неактивными до начала фазы S, когда их ингибитор Sic1 фосфорилируется под действием комплексов Cdc28 с циклинами фазы G1 и подвергается зависящему от убиквитина протеолизу. Белок Dbf4 очень нестабилен в течение всего клеточного цикла, особенно в начале фазы G1, когда его период полураспада равен 5 мин. Однако в начале фазы S его уровень и активность протеинкиназы Cdc7-Dbf4 достигают максимума. Таким образом, оба типа протеинкиназ становятся активными почти одновременно. Однако в активации ими ОНР проявляется определенная иерархия: Cdc7-Dbf4 действует после Cdc28-Clb. Одним из субстратов для Cdc28-Clb является белок Cdc6, который сыграл свою роль после погрузки комплекса МСМ. Этот белок после фосфорилирования покидает комплекс с ORC и подвергается протеолитической деградации. В освобождении связанного Cdc6 может играть роль гидролиз АТФ, связанного с белком Orc1. На освободившееся место вербуется новый важный компонент инициации репликации – белок Cdc45, который взаимодействует с белками МСМ. Для включения Cdc45 в предрепликативный комплекс требуется действие киназы Cdc7-Dbf4, которая связывается с белками МСМ и вызывает их фосфорилирование. Белки Mcm2-4 и Mcm6-7 являются субстратами для этой киназы in vitro. Фосфорилирование МСМ под действием Cdc7-Dbf4 сопровождается повышением чувствительности ДНК в области В2 ARS1 к KMnO4, взаимодействующему с онДНК. Это показало, что Cdc7-Dbf4 запускает переход ДНК геликазы МСМ в активное состояние (см. 2.1) и вызывает локальное расплетание ДНК в богатом А:Т сегменте ОНР. Связывание Cdc45 и модификация MCM приводят к образованию “преинициирующего комплекса”. В этот комплекс привлекаются также связывающий онДНК белок RPA и ДНК-полимераза a - праймаза. Вербовка этих белков критически зависит от присутствия в преинициирующем комплексе белка Cdc45, который может физически взаимодействовать с ними. В частности, белок Cdc45 связывается с субъединицей р70 комплекса Pola-праймаза (рис. 00). Киназа Cdc7-Dbf4 способна фосфорилировать и такие компоненты репликативного комплекса, как ДНК-полимераза a. Вербовка Pola-праймазы является первым этапом образования репликативных вилок на ARS.
Рис. 3.7. Этапы пути инициации репликации ДНК у дрожжей
Рис. 3.8. Схема участия белков МСМ, ДНК-полимеразы a - праймазы и белков Cdc45 и RPA в инициации репликации на эукариотической области ori
После образования двунаправленных репликативных вилок геликаза МСМ и белок Cdc45 выходят из контакта с комплексом ORC и перемещаются вместе с репликативной ДНК-полимеразой. На ДНК ARS остается только пострепликативный комплекс, содержащий ORC – как и в начале пути инициации репликации. Такие комплексы существуют в течение митоза и ранней фазы G1 на копиях ARS в обеих дочерних хромосомах.
Поиск по сайту: |