|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Погрузчики скользящего зажима
Кольца олигомерных форм белков DnaN и PCNA являются очень стабильными. Так, константа диссоциации димера DnaB не превышает 50 нМ, а период «полураспада» димерного кольца, надетого на ДНК, равен ~ 100 мин при 37о. Это означает, что спонтанная погрузка колец скользящих зажимов на кольцевую ДНК бактериальной хромосомы или на очень длинные эукариотические линейные хромосомы маловероятна. Для погрузки на ДНК необходимо временное размыкание колец хотя бы по одной из контактных поверхностей между субъединицами, чтобы через образующуюся щель внутрь кольца могла проникнуть ДНК. Этот процесс требует участия специальных молекулярных машин – погрузчиков зажима: g-комплекса бактериальной ДНК-полимеразы III или комплекса фактора репликации С (RFC) у эукариотов и археев. Эти комплексы содержат субъединицы, похожие на члены суперсемейства белков ААА+. Сокращенное обозначение ААА означает «АТФ-азы, ассоциированные с различными клеточными активностями».
Белки ААА+
Белки суперсемейства ААА+ действительно, участвуют в разнообразных клеточных процессах: 1) в АТФ-зависимом протеолизе короткоживущих и развернутых белков у бактерий (например, протеазы Lon и протеазные комплексы с участием субъединиц ClpA ClpX, гены которых входят в регулон теплового шока у E. coli); 2) в регуляции активности эукариотических протеасом 26S, каждая из регуляторных частиц которых содержит 6 разных, но близкородственных белков ААА; 3) в регуляции слияния мембран в процессах опосредованного везикулами транспорта (например, белок NSF/Sec18p) и передачи нервных импульсов; 4) в работе моторного белка динеина, переносящего грузы вдоль микротрубочек; 4) в генетической рекомбинации (например, бактериальный моторный белок RuvB, участвующий в миграции рекомбинационного холидеевского стыка по ДНК). Особую группу образуют белки ААА+, участвующие в репликации ДНК. Одни из них необходимы для сборки комплексов инициации репликации. К ним относятся бактериальный белок DnaA и эукариотические белки комплексов ORC и MCM (см. гл. 3). Вторую подгруппу составляют погрузчики скользящих зажимов ДНК-полимераз. Все белки ААА+ выполняют работу, сопряженную со связыванием и гидролизом АТФ, для выполнения их биологической функции, которая часто состоит в разборке белок-белковых и ДНК-белковых комплексов. Белки суперсемейства ААА имеет консервативный сегмент длиной около 220 остатков, который обычно называют модулем ААА, или модулем связывания нуклеотидов. Он состоит из доменов I и II, содержащих высококонсервативные мотивы аминокислотной последовательности. В домене I расположен активный центр связывания и гидролиза АТФ. Он содержит характерные для многих АТФаз мотивы Уокера (J. Walker) А и В. Мотив А с консенсусной последовательностью GxxxGKT, где х –произвольный остаток, образует в домене I так называемую Р-петлю, которая взаимодействует с необходимым для АТФазной активности катионом Mg2+ при участии остатка треонина и с b- и g-фосфатными группами АТФ при участии остатка лизина. Уокеровский мотив В (DExx) cодержит 2 смежные карбоксильные группы. Одна из них принадлежит остатку асп и входит к координационную сферу Mg2+, а вторая относится к остатку глу и считается каталитическим основанием АТФазы. Кроме этих необходимых для АТФазной активности участков, домены I и II содержат так называемые сенсорные мотивы 1 и 2, которые способны различать, какой нуклеотид (АТФ или АДФ) связан с активным центром белка ААА+. В таком распознавании важную роль играют консервативные остатки асн и тре сенсора-1 в домене I, способные образовывать водородную связь только с g-фосфатом АТФ. В сенсоре-2, находящемся с домене II, находится остаток арг, который электростатически взаимодействует с g-фосфатом АТФ. Гидролиз АТФ до АДФ устраняет узнаваемую сенсорами g-фосфатную группу. Это приводит к изменению конформации сенсора-2 и всего белка ААА+. Таким образом, белки ААА+ в комплексах с АТФ и АДФ могут находиться в разных конформационных состояниях. Изменение конформации этих белков может играть роль молекулярного переключателя. В этом отношении белки ААА+ похожи на маленькие связывающие гуаниновые нуклеотиды эукариотические мембранные белки типа Ras в цепи передачи сигнала, которые также находятся в разных конформационных состояниях: активном в комплексе с ГТФ и неактивном в комплексе с ГДФ. Дополнительный домен 3, прикрепленный к модулю ААА у некоторых белков ААА+, не консервативен у разных типов этих белков, и может, участвовать, например, в образовании их олигомеров. Все белки ААА+, для которых установлена 3-мерная структура, являются кольцевыми олигомерами. Их кольца чаще состоят из 6 и реже из 5 или 7 субъединиц. В мультимерных комплексах белков ААА+ сенсорные мотивы одного протомера иногда могут узнавать g-фосфатную группу АТФ, связанного с соседним протомером. Поэтому переход АТФ®АДФ в одной субъединице может сопровождаться изменением конформации смежных субъединиц.
g-Комплекс погрузчика зажима ДНК-полимеразы III E. coli
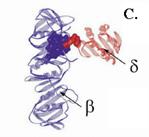
Из пяти разных субъединиц g-комплекса ДНК-полимеразы III E. coli только три (g, d и d’) существенны для действия погрузчика скользящего зажима. Рентгеноструктурный анализ субкомплекса gdd’ показал, что он содежит три протомера g и по одному протомеру d и d’. Данные, полученные при реконструкции этого субкомплекса in vitro, продемострировали, что любая из субъединиц g может быть заменена на более длинную субъединицу t, так что субкомлекс может содержать от 3 протомеров g до 3 протомеров t. Следовательно, уникальные для t С-концевые домены IV и V (см. 1.2.3) не мешают работе погрузчика. Субъединица g в g-комплексе наиболее похожа на белки семейства ААА+. Она имеет форму буквы С и состоит из 3 доменов (рис. 1.18, А). Типичный N-концевой домен, похожий по общей укладке на домен связывания АТФ в рекомбинационном белке RecA, содержит уокеровские домены А (GTRVGDKTS) и В (DEVH) и сенсор-1. Короткий соединительный домен II, состоящий из 4 a-спиралей, содержит сенсор-2 с типичным остатком арг. С-концевой домен III, имеется только у белков-погрузчиков в суперсемействе ААА+. Он состоит из нескольких a-спиралей и участвует в белок-белковых взаимодействиях. Субъединицы d и d’ по первичной аминокислотной последовательности имеют низкий уровень гомологии с g, причем субъединица d наименее консервативна. Тем не менее, рентгенгеноструктурный анализ индивидуальной субъединицы d’ и субъединицы d в составе субкомплекса g3dd’ показал, что по 3-мерной структуре эти компоненты субкомплекса очень похожи на белки ААА+ по общей укладке. Их молекулы имеют форму буквы С и содержат типичные для белков ААА+ пространственные домены I и II, а домены III, как и в субъединице g, состоят преиущественно из a-спиралей. С другой стороны, в доменах I субъединиц d и d’ отсутствуют компоненты активного АТФазного центра. Так, у белка d’ в Р-петле существенные остатки KT заменены на DD, в уокеровском мотиве В отсутствует один из кислых остатков, а в сенсоре-2 из домена II консервативный остаток арг заменен на гли. Еще более сильно изменена структура этих мотивов в субъединице d. Следовательно, эти субъединицы не могут связывать и гидролизовать АТФ. Такой активностью обладает только субъединица g, считающаяся мотором погрузчика скользящего зажима.
Рис. 1.1800. Структура субъединиц g (А) и d’ или d (В) комплекса погрузчика скользящего зажима ДНК-полимеразы III E.coli. BIE – участок взаимодействия с b-субъединицей, содержащийся только в субъединице d
Из всех индивидуальных компонентов субкомплекса gdd’ только свободная субъединица b с наибольшим сродством связывается с b-кольцом, надетым на ДНК, и может вызывать его освробождение, т.е. размыкать контакты между b-субъединицами в димере. Свободная субъединица g связывается с b-кольцом гораздо слабее и в 20 раз менее эффективно разгружает его с ДНК. Субъединица d’ не может вызывать разгрузку b-колец. Более того, связывание d’ с d ингибирует разгрузку b-кольца. Таким образом, характеристической способностью вызывать размыкание колец димерной формы b обладает только субъединица d. Ретнгеноструктурный анализ комплекса субъединиц d и b позволил установить молекулярный механизм размыкания d-кольца. Из трех доменов d-субъединицы только N-концевой домен I может контактировать с b. Специфический выступ на дистальном конце этого домена, напоминающий по форме треугольный клин и содержащий существенные гидрофобные остатки, внедряется в гидрофобную щель между доменами 2 и 3 на одной стороне внешней поверхности субъединицы b (рис. 1.19, С). Белок d взаимодействует не контактными поверхностями мономеров в димере b, а с консервативным участком, соотетствующим сайту взаимодействия b с a-субъединице ДНК-полимеразы III. Cравнение конфигурации мономера b-субъединицы в свободной форме или в форме открытого кольца после взаимодействия с d и в закрытой форме свободного b-кольца показало, что кривизна b-субъединицы в первом случае гораздо меньше, чем во втором. Это позволило предположить, что в закрытой форме кольца поверхность контакта между мономерами b находится в напряженном состоянии и что взаимодействие поверхности b-субъединицы с клином белка d вызывает изменение конформации этого мономера b, снимающим это напряжение и вызывающим образование щели на ближайшей поверхности контакта b-b. Размер этой щели достаточен для того, чтобы пропустить внутри кольца или из него онДНК, но не днДНК. Этот процесс идет в отсутствие АТФ, т.е. не требует затараты энергии. Таким образом, белок d действует как молекулярный клин, индуцирующий изменение конформации и вызывающий раскрывание b-кольца. В клетке E.coli содержится ~ 950 молекул d и только 140 молекул ее ингибитора – белка d’. Следовательно, около 800 молекул белка d находятся в свободном состоянии и могут участвовать в разгрузке с ДНК b-колец, покинутых ДНК-полимеразой III во время синтеза отстающей нити. Это способствует рециклированию b-колец, общее число которых на клетку составляет ~ 300 и гораздо меньше общего числа скользящих зажимов, требующихся для полной репликации всего генома E. coli (2000-4000). Несмотря на присутствие d-субъединицы в субкомплексе g3dd’, он не может в отсутствие АТФ вызывать размыкание b-колец, необходимое для их погрузки на ДНК. Однако в присутствии АТФ комплексы g3dd’ служат эффективными погрузчиками. Это позволило предположить, что в отсутствие АТФ в комплексе g3dd’ субъединица d’ маскирует N-концевой клин субъединицы d, не давая ему взаимодействовать с поверхностью b-кольца. В присутствии АТФ конформация всего комплекса изменяется, он переходит в более открытую форму, в которой субъединица d становится доступной для взаимодействия с b-кольцом. Изучение кристаллической структуры комплекса g3dd’ подтвердило эту гипотезу. В петамерном кольцевом комплексе g3dd’ индивидуальные субъединицы расположены по кругу в следующем порядке: …d’-g1-g2-g3-d… (рис. 1.19, А). С-концевые домены III всех субъединиц участвуют в их ассоциации в пентамер. Ассоциация обеспечивается встраиванием a-спирали первого партнера каждой пары в гидрофобную щель между двумя a-спиралями следующего партнера. По 3-мерной структуре комплекс g3dd’ похож на кисть правой руки, сложенную в щепоть (рис. 1.19, В). Соединенные С-концевые домены всех субъединиц соответствуют ладони, а домены I и II вместе взятые –пальцам (субъединица d’ – большой палец, три смежные субъединицы g1, g2 и g3 - указательный, средний и безымянный пальцы и субъединица d - мизинец).
Рис. 1.1900. Трехмерная структура комплекса погрузчика скользящего зажима ДНК-полимеразы III E. coli. А – вид сверху, В – вид сбоку (в неактивном состоянии в отсутствие связанного АТФ), С – взаимодействие N-концевой области субъединицы d с кольцом субъединицы b
Можно предположить, что в отсутствие АТФ комплекс g3dd’ находится в «закрытой» конфигурации, в которой кончики «пальцев» тесно прижаты друг к другу. В этом состоянии N-конец субъединицы d’ маскирует в субъединице d участок клина BIE, взаимодействующего с b-кольцом, и комплекс g3dd’ не может вызывать размыкание кольца. При взаимодействии с АТФ происходит постепенный переход комплекса в открытую конфигурацию, обусловленный изменением коформации всех субъединиц, за исключением d’, которая играет роль конформационно стабильного статора в погрузчике зажима. Три молекулы АТФ связываются только с 3 субъединицами g и, вероятно, в строго определенном порядке. Вначале АТФ взаимодействует только с субъединицей g1, т.к. только на стыке d’-g1 домен связывания АТФ в субъединицах g открыт для АТФ. Образование комплекса g1-АТФ вызывает характерные для белков ААА+ конформационные изменения на стыке g1-g2 при участии сенсорных доменов, открывающие для связывания АТФ домен I в g2 и т.д. Связывание АТФ с последней АТФазной субъединицей g3 вызывает изменение конформации в белке d, N-концевой домен которого спасается от секвестрирования смежной субъединицей d’. Это придает комплексу g3dd’ способность ассоциироваться с b-кольцом и раскрывать его по одной из контактных границ между мономерами. Таким образом, при раскрывании b-кольца АТФ необходим только для изменения конформации комплекса g3dd’. Эти события соответствуют переходам из состояния “a” в состояние “c” на общей схеме погрузки скользящего зажима на ДНК (рис. 1.20). Комплекс g3dd’ в отсутствие b-кольца имеет очень низкое сродство к ДНК независимо от присутствия АТФ. Однако в присутствии белка b он приобретает способность связываться с ДНК, предпочитая стыки затравка-матрица. По-видимому, после открывания b-кольца через щель между мономерами субъединицы b внутрь кольца пропускается нить онДНК на таких стыках, и комплекс g3dd’ прикрепляется к ДНК (cостояние “d”). Взаимодействие с ДНК по неустановленному механизму активирует АТФазую активность g-субъединиц и вызывает переход по меньшей мере двух из них в форму комплекса с АДФ. Смена партнера в АТФазных центрах этих субъединиц узнается их доменами сенсор-2 и приводит к развитию в комплексе g3dd’ обратных конформационных изменений, возращающих g3dd’ в исходную замкнутую конфигурацию. N-концевой сегмент BIE субъединицы d покидает поверхность b-кольца, которое смыкается на ДНК (состояние “e”). Комплекс g3dd’ освобождается от погруженного зажима, а последующее освобождение АДФ возвращает его в исходное состояние “a”. Цикл погрузки b-кольца на этом заканчивается (состояние “e”). Погрузка b-кольца сопровождается гидролизом АТФ, энергия которого израсходовалась на регенерацию замкнутой формы погрузчика.
Рис. 1.20. Последовательные этапы погрузки b-кольца скользящего зажима ДНК-полимеразы III E. coli на праймированную ДНК
Литература 1. Льюин Б. «Гены», М., Мир, 1987, с. 396-431. 2. Brush G.S., Kelly T.J. Mechanisms for replicating DNA // In “DNA Replication in Eukariotic Cells”, Cold Spring Harbor Lab. Press, Cold Spring Harbor, N.Y., 1996, pp. 1-43. 3. Alberts B. DNA replication and recombination // Nature, v. 421, 431-435, 2003. 4. Bollum F.J. Therminal deoxynucleotidyl transferase // In “The Enzymes”, v. 10, p.145-171, 1974. 5. Marians K. Prokaryotic DNA replication // Ann. Rev. Biochem., v. 61, 673-719, 1992. 6. Joyce C.M. Polymerase structures and function: variation on a theme? // J. Bacteriol., v. 177, 6321-6329, 1995. 7. Steitz T.A. DNA polymerases: structural diversity and common mechanisms // J. Biol. Chem., v. 274, 17398\5-17398, 1999. 8. Brautigam C.A., Steitz T.A. Structural and functional insights provided by crystal structures of DNA polymerases and their substrate complexes // Curr. Opin. Structural Biol., v. 8., 54-63, 1998. 9. Jäger J., Pata J.C. Setting a grip: polymerases and their substrate complexes // Curr. Opin. Structural Biol., v. 9., 21-28, 1999 10. Patel P.H., Suzuki M., Adman E., Shinkai A, Loeb L.A. Prokaryotic DNA polymerase I: evolution, structure, and “base flipping” mechanism for nucleotide selection // J. Mol. Biol., v. 308, 823-837, 2001 . 11. Михайлов В.С. ДНК-полимеразы эукариот // Мол. биол., т. 33, 567-580, 1999. 12. Stuckl M., Stagljar I., Jonsson Z.O., Hűbscher U. A coordinated interplay: proteins with multiple functions in DNA replication, DNA repair, cell cycle / checkpoint control, and transcription // Progr. Nucleic Acid Res. Mol. Biol., v. 65, 261-299, 2001. 13. Hűbscher U., Maga G., Spadari S. Eukaryotic DNA polymerases // Ann. Re. Biochem., v. 71, 000-000, 2002. 14. Budd M.E., Campbell J.I. Interrelationships between DNA repair and DNA replication // Mutat. Res., v. 451, 241-255, 2000 15. Foiani M., Lucchini G., Plevani P. The DNA polymerase a - primase complex couples DNA replication, cell cycle progression and DNA damage response // Trends Biochem. Sci., v. 22, 424-427, 1997 16. Burgers P.M.J. Eukaryotic DNA polymerases in DNA replication and repair // Chromosoma, v. 107, 218-227, 1998. 17. Burgers P.M.J., Koonin E.V. et al. Eukaryotic DNA polymerases: proposal for a revised nomenclature // J. Biol. Chem., v. 276, 43487-43490, 2001. 18. Aravind L., Koonin E.V. Phosphoesterase domains associated with DNA polymerases of .different origins // Nucl. Acids Res., v. 26, 3746-3752, 1998. 19. Cahn I.K.O., Ishino Y. Archaeal DNA replication: identifyng the pieces to solve a puzzle // Genetics, v. 152, 1249-1267, 1999. 20. Hingorani M.M., O’Donnell M. Sliding clamps: a (tail)ored fit // Curr. Biol., v. 10, R25-R29, 2000. 21. Tsurimoto T. PCNA, a multifunctional ring on DNA // Biochem. Biophys. Acta, v. 1443, 23-39, 1998. 22. Lopes de Saro F.J., O’Donnell M. Interaction of the b sliding clamp with MutS, ligase and DNA polymerase I // Proc. Nat. Acad. Sci. USA, v. 98, 8376-83802, 2001. 23. Dalrymple B.P., Kongsuwan K., Wijffels G., Dixon N.E., Jennins P.A. A universal protein-protein interaction motif in the eubacterial DNA replication and repair systems // Proc. Nat. Acad. Sci. USA, v. 98, 11627-11632, 2001. 24. Mossi R., Hűbscher U. Clamping down on clamps and clamp loader. The eukaryotic replication factor C // Eur. J. Biochem., v. 254, 209-216, 1998. 25. Trakselis M.A., Benkovic S.J. Intricacies in ATP-dependent clamp loading: variation across replication systems // Structure, v. 9, 999-1004, 2001. 26. Ellison V., Stillman B. Opening of the clamp: an intimate view of an ATP-driven biological machine // Cell, v. 106, 655-660, 2001. 27. Neuwald A.F., Aravind L., Spouge J.L., Koonin E.V. AAA+: a class of chaperone-like ATPases associated with assembly, operation, and disassembly of protein complexes // Genome Res., v. 9, 27-43, 1999. 28. Vale R.D. AAA proteins: lords of the ring // J. Cell Biol., v. 150, F13-F19, 2000.
Глава 2. Вспомогательные белки репликации ДНК ДНК-геликазы
Поиск по сайту: |