|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Сократительно-двигательный аппарат низших турбеллярий
Наиболее примитивные из турбеллярий – низшие Acoela – по общему уровню организации скорее приближаются к личинкам кишечнополостных, паренхимулам и планулам, нежели к взрослым Cnidaria. В частности, они характеризуются слабой степенью эпителизации своего фагоцитобласта, значительным развитием аморфной паренхимы и значительными формативными возможностями последней. В связи с этим наряду с эпидермальной мускулатурой все турбеллярии имеют хорошо развитую паренхимную, фагоцитобластическую мускулатуру, которую мы уже видели и у гребневиков. Эпидермальная мускулатура в простейшем случае состоит из миофибрилей, дифференцированных в толще эпидермиса. Они образуют 2 или 3 слоя; имеются наружные кольцевые и внутренние продольные волокна; иногда, кроме того, встречается и слой наружных продольных мышц (многие Anaperidae) или слой диагональных мышц. Таким образом, эпидермальная мускулатура Acoela образует сплошной мешок, состоящий из двух или трех слоев мышечных волокон и одевающий все тело, т.е. представляет собой один из вариантов кожно-мышечного мешка. С механической точки зрения кожно-мышечный мешок плоских червей очень напоминает то, что мы видим у гидр или актиний. Во всех случаях он образован слоями кольцевых и продольных мышц и заполнен жидкостью; главное отличие заключается в том, что у полипов – это жидкость, заполняющая их гастроваскулярную полость, а у плоских червей – собственные полужидкие ткани их тела. Однако при сокращении наружного мышечного мешка содержимое и здесь не поддается сжатию, давление внутри тела повышается и благодаря этому различия в тонусе отдельных групп кожных мышц вызывают изменения формы тела. Многие турбеллярий могут целиком вытягиваться в ниточку, сокращаться в шарик или вытягивать передний или задний конец тела. При этом все их внутренние органы, в зависимости от консистенции своих тканей, претерпевают либо сильнейшие временные деформации, либо перемещения, но и то и другое – без вреда для себя. Тем не менее необходимость выносить подобные воздействия, несомненно, отражается на всей организации животного и, вероятно, препятствует достижению органами высокого функционального совершенства. С другой стороны, «мезенхимная» организация, при которой весь кожно-мышечный мешок заполнен мезенхимой с заключенными в ней органами, представляет и некоторые конструктивные преимущества; одним из таких преимуществ является полная свобода возникновения мышечных волокон или пучков, пересекающих тело в любом направлении и появляющихся (в процессе эволюции) всюду, где в них возникает надобность. В простейшем случае паренхимные мышцы Acoela бывают представлены отдельными волокнами, которые либо беспорядочно идут по всем направлениям, либо более или менее придерживаются трех основных направлений: продольного, поперечного и дорсовентрального (рис. 128, А).При этом за счет продольных мышц очень часто обособляются системы ретракторов переднего и заднего концов тела, а за счет дорсо-вентральных мышц – ретракторы глотки. Даже у форм, имеющих обособленный пищеварительный синцитий, мышечные клетки или целые их пучки свободно пронизывают этот последний; мало того, и за пределами Acoela мы встречаем случаи, когда дорсовентральные мышцы тела пронизывают амебоидный эпителий кишечника (рис. 128, Б).По сравнению с высокополяризованной структурой и упорядоченными слоями мышечных отростков гидроидов, Acoela с их паренхимой, во всех направлениях пронизанной диффузноразбросанными мышечными волокнами, представляют, несомненно, более примитивное состояние, но более чреватое разнообразными возможностями прогрессивного развития. Кроме функции общей сократимости тела, мускулатура низших турбеллярий обычно обслуживает рот и совокупительный аппарат. Последнего у Xenoturbella нет, и мы встречаем у нее только те первичные функции сократительно-мышечного аппарата, которые видели и у кишечнополостных: Что касается функции передвижения, она у большинства Acoela всецело осуществляется ресничками кожного эпителия, но во многих случаях, особенно у более крупных (Апарегик, Amphiscolops) или сильно вытянутых форм (Proporus), известное локомоторное значение получает и мускулатура тела.
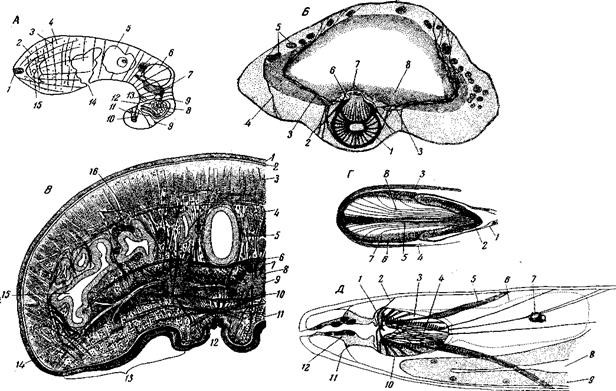
Рис. 128. Строение сократимого аппарата плоских червей А – Pseudoconvoluta flava (Acoela), медиальный разрез, показывающий схему расположения паренхимной мускулатуры: 1 – лобные железы; 2 – ретракторы переднего конца; 3 – дорсо-вентральные мышечные волокна; 4 – поперечные мышечные волокна; 5 – медиально расположенный овоцит; 6 – совокупительная сумка; 7 – потоки спермы, идущие к совокупительному органу (8); 9 – ретракторы совокупительного органа; 10 – железистый шип совокупительного аппарата; 11 – его протракторы; 12 – мужское половое отверстие; 13 – протракторы совокупительного органа; 14 – рот с зачаточной глоткой; 15 – мозг со статоцистом (оригинал); Б – Phaenocora potycirra (Rhabdoeoela Typhloplanoidea), поперечный разрез, показывающий олигомеризованный тип мускулатуры прямокишечных турбеллярий: 1 – мужской совокупительный орган; 2 – его вентральные ретракторы; 3 – его дорсальные ретракторы, пронизывающие стенку кишечника; 4 – дорсовентральные мышцы; 5 – разветвление желточника; 6 – мужской половой канал; 7 – протракторы совокупительного органа; 8 – эпителий кишечника (по Беклемишеву); В – Arliodactylus speciosus (Triclada Terricola), поперечный срез в области присасывательной ямки: 1 – эпидермис; 2 – его пограничная пластинка; 3 – продольные мыщцы паренхимы спинной стороны; 4 – фолликулы желточников; 5 – дорсовентральные мышцы: 6 – поперечные мышцы; 7 - железистые клетки, протоки которых открываются на ползательных полозьях; 8 – яйцевод; 9 – стволы глубокого внутрипаренхимного нервного сплетения; 10 – поперечные мышцы вентральной стороны; 11 – железы присасывательной ямки; 12 – ползательные полозья; 13 – присасывательная ямка; 14 – продольные мышцы; 15 – ветви кишечника; 16 – поперечные мышцы спинной стороны (по Graff); Г – хоботок Phonorhynchus mamertinus (Rhabdocoela Polycystididae) сагиттальном разрезе: 1 – хоботное влагалище; 2 – клейкий эпителий на конце хоботка; 3 – спинной протрактор; 4 – брюшной протрактор; 5 – протоки железистых клеток, открывающиеся на конце хоботка; 6 – пограничная пластинка хоботка; 7 – кольцевая мускулатура хоботка; 8 – продольная мускулатура хоботка (по J. Meixner); Д – хоботок Gnaihortnjnchus conocaudatus (Rhabdocoela Gnathorhynchidae) схема строения переднего конца тела, вид с левой стороны: 1 – крючья хоботка; 2 – передний ретрактор хоботка; 3 – продольный мышечный валик, служащий опорой крючку; 4 – внутренние ретракторы хоботка; 5 – наружные ретракторы хоботка; 6 – контуры мозга (пунктир); 7 – глаз; 8 – полость кишечника; 9 – его эпителий; 10 – мускулы-аддукторы крючьев; 11 – дилататоры хоботного влагалища; 12 – эпителий хоботного влагалища (по J. Meixner) Вытянутые формы при движении помогают себе змеевидным изгибанием тела. Сократимость тела и способность сильно менять форму очень помогают протискиванию в узких пространствах, что особенно существенно для живущих в толще грунта форм (например, Convoluta roskoffensis). Вообще при наличии общей сократимости тела и движения при помощи ресниц по поверхности субстрата или внутри субстрата участие мускулатуры в передвижении рождается само собой и на каждом шагу. Поэтому не удивительно, что все крупные турбеллярии в той или иной мере переходят к передвижению при помощи мышц; участие в ползании сохраняют только ресницы брюшной стороны (подошвы); работа остальных ресниц имеет у них только гидрокинетическое значение. В сущности, лишь возможность использовать для передвижения мускулатуру и позволила многим турбелляриям достичь крупных размеров, при которых уже невозможно передвижение при помощи одних ресниц. У турбеллярии встречаются 4 типа передвижения при помощи мускулатуры: 1) ползание путем перистальтики всего тела; 2) ползание при помощи перистальтических волн, проходящих по обращенной к субстрату стороне тела; последняя в таком случае отличается особенно сильным развитием мускулатуры, и в особенности дорсовентральной, и образует нечто
Рис. 129. Наземная планария Rhynchodemus bilineatus (Turbellaria, Triclada), ползущая при помощи перистальтики своей ползательной
вроде ползательной подошвы; иногда волны перистальтики на подошве появляются в виде настоящих поперечных валиков («м и о п о д и й»), так что ползущая планария касается субстрата только гребнями этих валиков (рис. 129), как, например, Rhynchodemus bilineatus (Triclada Terricola). При передвижении волн спереди назад вдоль подошвы животное катится на них, как на тракторных гусеницах; 3) движение при помощи змеевидных извиваний тела, свойственное вытянутым формам; при относительно слабой мускулатуре и значительных размерах тела мы видим змеевидное ползание, наиболее выраженное у Rhynchoscolex (Notandropora); при сильной продольной мускулатуре оно переходит в плавание; зачаточную форму такого плавания мы видим у Promonotus (Alloeocoela Crossocoela); при слабой мускулатуре и малых размерах тела сильно вытянутые формы плавают при помощи ресниц, но с участием изгибаний тела; 4) плавание при помощи сгибания и разгибания придатков тела. Среди турбеллярии этот способ передвижения в зачаточном виде свойствен многим поликладам, имеющим обособленные боковые края тела, играющие роль примитивных плавников. Развитие хватательно-глотательной функции выражается у высших турбеллярии прежде всего в создании разнообразных и часто сложных типов глотки. Сильная мускулатура и совершенство глотки позволяют турбелляриям овладевать добычей несмотря на слабость остальной мускулатуры тела. Хватательные щупальца, свойственные кишечнополостным, у турбеллярий полностью отсутствуют. Мозговые щупальца поликлад, быть может гомологичные аборальным щупальцам гребневиков и наркомедуз, сохранили только чувствительную функцию и полностью утратили хватательную. В качестве дополнительных органов овладевания добычей могут быть приспособлены края тела или его передний конец. Хватательные приспособления переднего конца тела в примитивных случаях выражаются, как мы видели, в присутствии клейких желез и мускулов-ретракторов. В подотряде Kalyptorhynchia (отряд Rhabdocoela) на переднем конце тела находится сложно дифференцированный хоботок, снабженный мышечным влагалищем, в которое он может втягиваться при помощи мускула-ретрактора, идущего от вершины хоботка ко дну влагалища; выпячивается хоботок благодаря сокращению стенки влагалища; снаружи к влагалищу прикрепляются мышцы, двигающие хоботок в целом, – ретракторы и протракторы. Хоботок может быть не расщеплен и снабжен на конце клейкими железками (Polycystididae и др.) (рис. 128, Г)либо раздвоен и иногда снабжен кутикулярными щипчиками (Schizorhynchidae, Gnathorhynchidae, рис. 128, Д). Хоботок Kalyptorhynchia расположен строго на переднем конце тела и потому его можно было бы рассматривать как видоизменение последнего. Однако в семействе Trigonostomidae сходный, хотя и лишенный влагалища хоботок образуется субтерминально, на брюшной стороне головы. У паразитических плоских червей хоботки, сходные по своему устройству с хоботком калипторинхий, но снабженные шипиками или крючочками и служащие органами прикрепления к хозяину, возникают в довольно различном числе и расположении. Многие циклофиллиды, например Taenia solium, имеют на сколексе единственный апикально расположенный хоботок; все тетраринхиды имеют по четыре таких хоботка, расположенных на сколексе радиально-симметрично (рис. 130, В). Дигенетический сосальщик Rhopalies имеет пару втяжных хоботков, расположенных по бокам ротовой присоски. Все это не позволяет считать хватательные и прикрепительные хоботки плоских червей видоизменением переднего конца тела животного; скорее в них можно видеть органы, возникающие путем местной перестройки покровов и мускулатуры и не обязательно на апикальном конце тела. Двигательный аппарат паразитических плоских червей, в принципе сходный с таковым турбеллярий, характеризуется, однако, сильным развитием прикрепительных органов. У Digenea обычно имеются две присоски – ротовая и брюшная, у Monogenea и их личинок задний конец тела обособлен в виде церкомера, несущего сложный прикрепительный аппарат из крючьев и присосок с соответствующей мускулатурой. В видоизмененном виде церкомер сохраняется и в развитии ленточных червей. Сколекс цестод также несет присоски и другие прикрепительные органы.
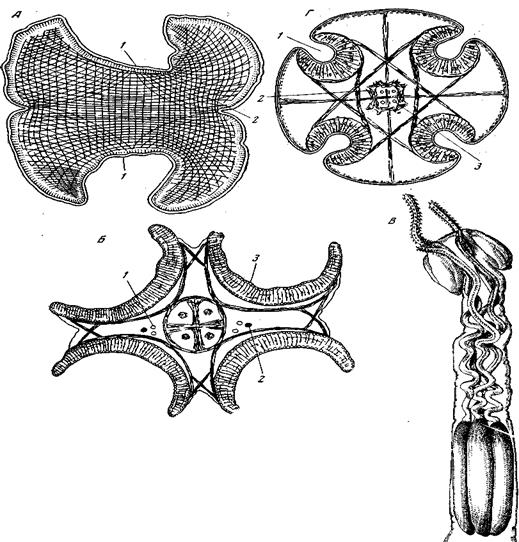
А – поперечный разрез через сколекс Pseudophyllidea с двумя присасывательными ямками: 1 – присасывательная ямка (bothriurn); 2 – мышечные волокна; Б –поперечный разрез через сколекс Tetraphyllidea с 4 ботриями: 1 – разрезы нефридиев; 2 – рзрезы нервных стволов; 3 – ботридии (из Hyman); В – сколекс Tetrarhynchus (Tetrarhynchidea); на переднем крае каждой из четырех присасывательных ямок открывается влагалище одного из четырех длинных хоботков; хоботок вооружен крючьями и впячивается в узкую часть влагалища при помощи длинного ретрактора; выворачивание хоботка совершается при помощи сокращения мощной мускулатуры проксимальной расширенной части влагалища; Г – поперечный разрез через сколекс Nematobothrium (Cyclophyllidae) с 4 присосками (acethabola) и двулучевым расположением мускулатуры: 1 – присоски; 2 – нервный аппарат сколекса; 3 – разрезы нефридиев (по Futirmann)
Поиск по сайту: |