|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Половой аппарат членистоногих и онихофор⇐ ПредыдущаяСтр 34 из 34
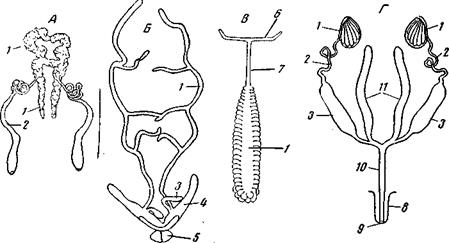
У всех членистоногих и онихофор, относительно которых эмбриология дает нам возможность судить, стенки гонад представляют участки стенок целома, и половые протоки, по крайней мере в своей проксимальной части, образованы целомодуктами. Далее, во всех случаях целомодукты прирастают к гонадам и вступают с ними, таким образом, в непосредственную связь, отсутствующую у аннелидного прототипа (рис. 173 и 174).
Рис. 173. Мужской половой аппарат членистоногих А – Palaemon (Crustacea Decapoda) (из Lang); Б – Scorpio maurus (Scorpionoidea) (по Павловскому); В – Glomeris marginata (Diplopoda) (no Pabre); Г – схема мужского полового аппарата насекомых (из Snodgrass); 1 – семенники; 2 – семевыносящий проток; Но в остальном наблюдается большое разнообразие: различны бывают и отделы целома (спинной, брюшной), за счет которых образуются гонады, и номера сомитов, в которых образуются гонады и их протоки; наконец, различным оказывается и происхождение полости гонад. Это многообразие говорит, скорее всего, о том, что в эволюции членистоногих процесс редукции целома и связанной с ним реконструкции полового аппарата протекал независимо в различных группах и принимал в разных группах разное течение.
Рис. 174. Женский половой аппарат членистоногих А – Mysis oculata relicta (Crustacea Mysidacea); 1 – зачатковая зона яичника;
Как гонады, так и целомодукты полихет парны и метамерны и у примитивных форм имеются во многих сегментах тела. Из современных членистоногих метамерно повторяющимися парными половыми отверстиями обладают только Pantopoda: в примитивных случаях и у самцов, и у самок они имеются на всех четырех парах ног. Эволюция идет в сторону олигомеризации числа отверстий, причем самцы опережают в этом отношении самок. Исчезновение отверстий идет спереди назад. Процесс совершается в нескольких стволах пантопод независимо и параллельно. Все остальные членистоногие сохраняют всего одну пару гонодуктов и одно, чаще непарное половое отверстие. Примитивная парность половых отверстий, кроме пантопод, сохранилась у большинства ракообразных, у Xiphosura, Diplopoda, Pauropoda и среди насекомых – у поденок (отряд Ephemeroidea) в обоих полах, у Protuга и некоторых уховерток – только у самцов. Форма и строение гонад и дифференцировка половых протоков членистоногих чрезвычайно разнообразны и в пределах отдельных классов представляют значительный морфологический интерес, но рассмотрение их выходит за объем настоящей книги. Отметим лишь некоторые особенности в эволюциии совокупительного аппарата членистоногих. Членистоногие выводятся из низших аннелид, т.е. из животных, обладающих наружным оплодотворением и лишенных совокупительных органов. Ввиду этого последние возникали у членистоногих независимо от их возникновения во всех нами выше рассмотренных группах. При этом чрезвычайное разнообразие совокупительных приспособлений у членистоногих показывает, что они возникали независимо в различных стволах этого подтипа. Среди хелицеровых мечехвосты имеют еще наружное оплодотворение: самка откладывает яйца в ямку, вырытую в песке на берегу моря, а самец поливает их спермой. Точно так же наружное оплодотворение имеет место и у пантопод: самец и самка прикладываются друг к другу брюшными сторонами (псевдокопуляция), самка выпускает яйца, самец – сперму; оплодотворенные яйца, слипшиеся между собой, наматываются на яйценосные ножки самца, где и вынашиваются. Из ракообразных некоторые совершенно лишены мужских совокупительных органов и при копуляции обе особи просто прикладываются друг к другу половыми отверстиями. У других имеется настоящий пенис, как, например, у Cirripedia, где он образован задним концом брюшка, а также у Priapion (Entoniscidae, отряд Isopoda), где он представляет вырост 7-го сегмента груди. У речного рака (Astacus) в совокупительные органы превращены I плеоподы, непосредственно примыкающие к половому отверстию, палочковидные и снабженные желобками. Оплодотворение яиц совершается вне половых путей матери, так как сперма при копуляции приклеивается к брюшной поверхности 8-го грудного сегмента самки. При откладке яиц самка загибает брюшко, прикрывая брюшную сторону груди, и выделяемый яйцеводами секрет растворяет секрет, склеивающий сперму; последняя освобождается и оплодотворяет выходящие из полового отверстия яйца. Этот своеобразный механизм оплодотворения, свойственный многим Масгага, очевидно, выработан в пределах отряда Decapoda и представляет видоизменение и осложнение наружного оплодотворения. У высших Decapoda (Brachiura) сперма при копуляции вводится прямо в женские половые пути. Среди Copepoda у Centropagidae левая 5-я грудная нога самца превращена в щипчики, обе половины которых образованы ее экзоподитом и эндоподитом. При помощи этих щипчиков самец захватывает из собственного полового отверстия сперматофор и переносит его в половое отверстие самки. Правая 5-я нога самца – крючкообразная и служит для придерживания самки. Этот тип мужского совокупительного органа отличается от того, что мы только что видели у Astacus, и принципиально сходен с тем, что имеет место у иксодид или бихорок, с той разницей, что у Centropagidae служащие для перенесения сперматофора конечности специально изменены для этой функции. Самки Gopepoda имеют самостоятельно открывающиеся наружу семеприемники, соединенные с терминальным отделом яичника тонкими протоками. Этим они напоминают самок пауков. Аналогичные женские совокупительные приспособления мы встречаем у многих плоских червей, пиявок и улиток. За последние 15–20 лет накопилось много данных о способах оплодотворения у наземных членистоногих, что позволило М.С. Гилярову воссоздать общую картину эволюции этого процесса. Переход членистоногих из воды на сушу совершился в основном через жизнь в почве, лесной подстилке и других защищенных местообитаниях. Оказывается, что многим, если не всем, первично-почвенным членистоногим свойствен один общий тип наружно-внутреннего оплодотворения. В одном из наиболее примитивных случаев самец приклеивает к частицам почвы маленькие булавовидные сперматофоры (клещи: Oribatei; насекомые: Gampodea) и делает это большей частью независимо от присутствия самки. Самка, случайно набредая на сперматофор, обследует его и затем «заглатывает» его своим половым отверстием. Или же самец натягивает в скважинах почвы нити секрета и помещает на них незащищенные капли спермы; самка, наткнувшись на нить, доходит по ней до спермы и также поглощает ее половым отверстием. Такую картину мы видим у многоножек. Самцы Pselaphognatha обладают органом, который обычно обозначается как пенис, но так как он служит не для введения спермы в тело самки, а для помещения ее на натянутую самцом нить, Гиляров его обозначает как сперматопозитор. Сперматопозитор свойствен и клещам-орибатидам. От описанного прототипа поведения идет множество рядов развития, во многом параллельных между собою. Так, самцы скорпионов и лжескорпионов прикрепляют сперматофоры к субстрату, у некоторых лжескорпионов еще независимо от присутствия самки, у большинства изученных лжескорпионов и всех изученных скорпионов, – во время брачной игры. В последнем случае самец чаще всего подтаскивает самку к сперматофору, который она и поглощает половым отверстием. У скорпионов при прикреплении сперматофора самцом и ощупывании его самкой большую роль играют гребни. Сходные типы оплодотворения имеют Scutigera и Sminthuridae (Gollembola). Сперматопозитор, возникающий среди акариформных клещей, например у Oribatei, у родственных им Acaridia используется как пенис; они перешли к внутреннему оплодотворению. Кроме части клещей, внутреннее оплодотворение среди паукообразных доказано для Opiliones подотряда Palpatores, у которых самец имеет длинный пенис и самка длинный яйцеклад, образованные за счет стенок эктодермального полового атрия. Другое направление эволюции намечается у Machilis germanica: самец, встретив самку, прикрепляет к субстрату нить и при помощи сперма-топозитора помещает на нее капли спермы; самка становится параллельно нити, а самец антенной собирает сперму и наносит в область полового отверстия самки – первый шаг в использовании для копуляции конечностей, не связанных непосредственно с половым отверстием (в данном случае – антенны). У насекомых этот принцип не получил большого развития, но в других группах членистоногих он играет значительную роль. Так, у сольпуг самец в присутствии самки выделяет комок спермы на землю, подбирает его хелицерами и при их помощи переносит в половое отверстие самки. У клещей отряда Parasitiformes оплодотворение совершается при помощи сперматофора, который самец берет хелицерами из собственного полового отверстия и переносит в половое отверстие самки. Хелицеры самца несколько видоизменены для этой цели. Пауки (Araneina) выводят сперму не на землю, а в сплетенную для этого предбрачную паутиновую сетку, откуда забирают ее в полость сложно видоизмененного последнего членика педипальп и затем при встрече с самкой, вводят ее в специальные семеприемники этой последней. Семеприемники чаще всего открываются протоками непосредственно наружу и, кроме того, соединены тонкими duct us spermatici с непарным яйцеводом (рис. 174, В), У самцов Ricinulei приспособлены для копуляции (превращены в гоноподии) ходильные ноги 3-й пары. Точно так же при помощи конечностей переносит сперму и большинство Diplopoda. Opisthandria осуществляют перенос ее из мужских отверстий в женские (и те и другие у них парные) при помощи мандибул и, таким образом, стоят в этом отношении на уровне паразитиформных клещей. У Proterandria одна, а иногда и обе пары конечностей 7-го диплосегмента превращены в гоноподии, часто весьма сложно устроенные Наружные половые органы Thysanura в обоих полах устроены довольно сходно. У самки они представляют яйцеклад (ovipositor), образованный гонапофизами 8 и 9-го сегментов брюшка. Самец имеет непарный сперматопозитор и по бокам его пару парамер; гомологии их не вполне установлены, но вероятно и тот, и другие являются производными гонапофиз 9-го сегмента. Высшие насекомые (Pterygota) имеют внутреннее оплодотворение, но ряд переходных форм указывает, что оно происходит от сперматофорного наружно-внутреннего. Сперматофоры сохраняются у насекомых с большим упорством и при чисто внутреннем оплодотворении, и исчезают только в высших отрядах: так, их нет у Hymenoptera и у большинства Coleoptera и Diptera. Из двукрылых они еще сохранились, например, у Culicoides. Половые придатки Pterygota гомологичны таковым Thysanura. Самки многих отрядов имеют яйцеклад (у части Hymenoptera превращенный в жало). Сперматопозитор Thysanura, путем смены функций, сделался пенисом; по бокам его, за счет коксоподитов и стилей 9-го сегмента, образуются хватательные придатки, служащие для придерживания самки при копуляции. Поденки и часть уховерток имеют парные пенисы. В различных отрядах насекомых копулятивные органы как мужские, так и женские представляют бесчисленные видоизменения, рассмотрение которых завело бы нас далеко.
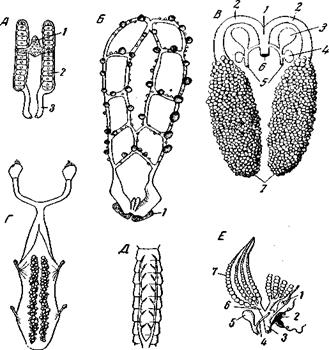
Рис. 175. Наружно-внутреннее оплодотворение низших наземных членистоногих А – капли спермы на паутинных нитях Polyxenus lagurus (Diplopoda, Pselaphognatha);
Итак типы копуляции и копулятивных органов членистоногих представляют множество рядов развития, частью параллельных, частью дивергирующих. В начале этих рядов лежит наружное оплодотворение, еще сохранившееся до наших дней у Xiphosura и Pantopoda. У почвенных Che-licerata и Atelocerata (pиc, 175) господствует наружно-внутреннее оплодотворение, весьма сходное с оплодотворением живущего в морском песке Protodrilus (хотя и обеспеченное совершенно другими исполнительными органами). Не исключено, что предки хотя бы части наземных членистоногих приобрели такой способ оплодотворения еще в море; в пользу этого говорит наличие гребней у силурского морского скорпиона Palaeophonus. Все способы оплодотворения других наземных членистоногих выводятся из этого типа, и для их осуществления в различных рядах возникли различные копулятивные органы. При этом мужские копулятивные органы возникают чаще всего по одному из следующих трех типов, которые мы видим и у ракообразных: 1) за счет разрастания и выпячивания краев полового отверстия или стенок полового атрия (например, у Орilinеs); 2) за счет конечностей, непосредственно примыкающих к половому отверстию (например, у большинства Insecta Pterygota) и 3) за счет конечностей, не примыкающих к половому отверстию (например, у сольпуг, пауков, большинства Diplopoda и пр.). В двух первых случаях, но не в третьем, настоящие копулятивные органы, по-видимому, очень часто возникали путем смены функции из сперматопозиторов (Oribatei – Acaridia, Thysanura – Pterygota). Способы копуляции и копулятивные органы ракообразных еще нуждаются в приведении их в систему. Onychophora по способу оплодотворения резко отличаются от членистоногих; самец втыкает сперматофор в кожу самки и оплодотворение сводится к гиподермической импрегнации, как у части пиявок.
Вопросы для самоконтроля 1. На какие типы подразделяются многоклеточные в зависимости от времени обособления половых клеток? 2. Что такое половой аппарат? 3. Какие способы встречи яйца и сперматозоидов существуют у беспозвоночных животных? 4. Что входит в состав полового аппарата? 5. Эволюция полового аппарата губок. 6. Эволюция полового аппарата кишечнополостных. 7. Строение полового аппарата турбеллярий. 8. Строение полового аппарата цестод. 9. Особенности строения половой системы сосальщиков. 10. Формирование полового аппарата аннелид. 11. Как устроен половой аппарат моллюсков? 12. Строение мужского полового аппарата членистоногих. 13. Строение женского полового аппарата членистоногих. ЛИТЕРАТУРА 1. Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных. – М., 1964, т. 1. – 432 с., т. 2. – 446 с. 2. Беклемишев К.В. Зоология беспозвоночных: Курс лекций. М., 1979. – 188 с. 3. Гиляров М.С. Биологический энциклопедический словарь. – М., 1986. – 828 с. 4. Догель В.А. Зоология беспозвоночных. – М., 1981. – 606 с. 5. Захваткин А.А. Сравнительная эмбриология низших беспозвоночных животных. – М., 1949. – 395 с. 6. Иванов А.В. Происхождение многоклеточных животных. – Л., 1968. – 288 с. 7. Иванова-Казас О.М. Сравнительная эмбриология беспозвоночных животных. Простейшие и низшие многоклеточные. – Новосибирск, 1975. – 372 с. 8. Клюге Н.Ю. Современная систематика насекомых. – СПб., 2000. – 332 с. 9. Лопатин И.К. Функциональная зоология. – Мн., 1998. – 230 с. 10. Мазохин – Поршняков Г.А. Зрение насекомых. – М., 1965. – 277 с. 11. Просир Л. Сравнительная физиология животных. – М. т. 2., 1977. – 571 с. 12. Райков И.Б. Ядро простейших. – Л., 1978. – 328 с. 13. Серавин Л.Н. Простейшие… Что это такое? – Л., 1984. – 174 с. 14. Шарова И.Х. Зоология беспозвоночных. – М., 1999. – 592 с. 15. Царичкова Д.Б. Методические указания к разделу «Сравнительная анатомия низших беспозвоночных». – Киев, 1985. – 22 с. 16. Царичкова Д.Б. Физиология нервной системы и органов чувств насекомых. – Киев, 1978. – 48 с.
[1] Ретинулой называется совокупность зрительных клеток одного омматидия.
[2] Термин «целомодукт» предложен Ray lankester (1900).
Поиск по сайту: |