|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Кровеносный аппарат членистоногих и онихофор
Совершенно иную судьбу кровеносного аппарата наблюдаем мы у членистоногих и близких к ним в этом отношении онихофор. Членистоногие выводятся из аннелидообразного прототипа с хорошо развитой замкнутой кровеносной системой; замкнутое кровообращение они утратили все, и многие из них представляют различные степени упрощения, а иногда и редукции кровеносного аппарата. Судьба кровеносного аппарата членистоногих тесно связана с судьбой целома. С распадом стенок целома за их счет образуется, помимо мускулатуры, рыхлая целенхимная ткань, выполняющая промежутки между органами, бедная промежуточным веществом и содержащая многочисленные заполненные гемолимфой пространства. В этом отношении гистологическое строение членистоногих больше напоминает моллюсков, нежели аннелид. В силу этого у них отсутствует и замкнутая кровеносная система аннелид, проходящая в толще системы пограничных пластинок. Из кровеносного аппарата аннелид членистоногие сохранили магистральные пути: спинной сосуд, иногда брюшной сосуд и боковые дуги; в то же время все они полностью утратили капилляры и мелкие сосуды. Все крупные сосуды приобретают значение артерий, по которым кровь расходится из спинного сосуда или возникшего за его счет сердца. Из последних разветвлений этих сосудов кровь изливается в лакунарные пространства, по которым и возвращается к сердцу. Лишь у немногих членистоногих образуются венозные стволы, несомненно вторичного происхождения, представляющие такую же канализацию лакунарных пространств, как и венозные стволы моллюсков. Мнение, что для всех членистоногих характерна тенденция к постепенной редукции всего кровеносного аппарата, справедливо лишь для насекомых и для карликовых форм из других классов. Как и моллюскам, членистоногим свойственна корреляция между величиной тела и степенью развития кровеносного аппарата: чем меньше членистоногое, тем меньше разветвлена его артериальная система. У мелких членистоногих от всего кровеносного аппарата остается только сердце, как, например, у Gladocera, у Gamasides среди Parasitiformes, у Calanidae среди Copepoda (рис. 156, В), или даже исчезают всякие следы кровеносного аппарата, как у большинства Copepoda, Ostracoda и клещей. Слабое развитие или полная редукция кровеносного аппарата у мелких членистоногих, как и у мелких моллюсков, связаны с тем, что у таких форм все органы лежат очень близко друг от друга и функция распределения может полностью осуществляться при посредстве лакунарных полостей и того передвижения жидкостей в этих последних, которое достигается при движении тела. Заметим, что у мелких аннелид кровеносная система всегда остается замкнутой, а в соответствии с малой величиной уменьшается лишь степень ветвления сосудов. Спинной сосуд членистоногих по способу своего развития гомологичен, как мы видели, спинному сосуду аннелид. В наиболее примитивных случаях он тянется на всем или почти на всем протяжении туловища. Таковы Phyllopoda и Stomatopoda среди ракообразных, скорпионы – среди хелицеровых, Chilopoda и Diplopoda – среди многоножек, Blattodea – среди насекомых. Точно так же вдоль всего туловища тянется спинной сосуд и у Onychophora. Спинной сосуд Onychophora или Anostraca на всем своем протяжении сократим и снабжен метамерно расположенными парными остиями. У Branchipus их имеется полное число – 18 пар. Остии представляют снабженные клапанами входные отверстия, через которые кровь засасывается в сосуд. Сократимый спинной сосуд, снабженный остиями, носит у членистоногих название сердца.
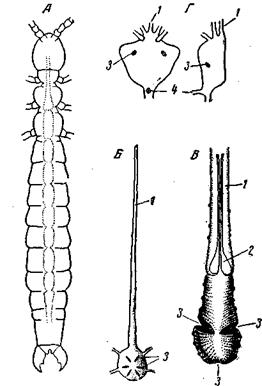
Рис. 156. Строение сердца некоторых членистоногих А – Japix (Campodeoidea) (по Grasse); Б– Наеmatopinus (Anoplura) (из Weber);
У многих форм по ходу его располагаются метамерные клапаны, допускающие движение крови лишь в одном направлении – сзади наперед. В ряде других случаев происходит дифференцировка спинного сосуда: передний конец его утрачивает мускулатуру, остии и клапаны и превращается, таким образом, в переднюю аорту. Иногда таким же путем образуется и задняя аорта, как у части моллюсков. Так возникает гетерономное строение спинного сосуда. Его сократимый участок, или сердце, в различных классах и отрядах располагается в различных частях тела (в разных тагмах). Иногда сердце подвергается значительному укорочению – главным образом у ракообразных, но также и у отдельных хелицеровых (лжескорпионы, сенокосцы, клещи) и насекомых (например, вши; рис. 156, Б). У раков укорочение сердца происходит не только при редукции кровеносного аппарата (мелкие Entomostraca), но также и при высоком уровне организации этого последнего. Так, например, Decapoda (рис. 156, Г)имеют укороченное, мешковидное сердце с 3 парами остии при весьма совершенном кровеносном аппарате. Здесь укорочение сердца является крайним проявлением дифференцировки спинного сосуда и централизации пропульсаторной функции, т.е., несомненно, прогрессивным признаком.
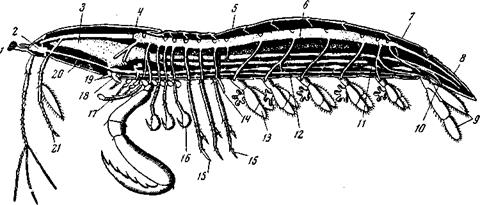
Рис. 157. Схема строения Squilla (Stomalopoda), самец 1 – глаза; 2 – мозг; 3 – жевательный желудок; 4 – саrарах; 5 – первый членик брюшка; 6 – сердце, с отходящими от него передней и задней аортой и 15-ю парами артерий, а также со стигмами; 7 – последний сегмент брюшка; 8 – тельсон; 9 – уроподы; 10 – анус; 11 – брюшная нервная цепочка; 12 – субневральная артерия;
Укорочение и дифференцировка спинного сосуда является основной прогрессивной тенденцией в эволюции кровеносного аппарата членистоногих. Явление это представляет известную формальную аналогию с укорочением у них же брюшной нервной цепочки и также связано с нарастанием гетерономности всей метамерии тела. Брюшной сосуд имеется у немногих артропод: у многоножек, скорпионов, мечехвостов и у ряда высших раков (рис. 157, 158). У скорпионов брюшной сосуд лежит между нервной цепочкой и кишечником, образуя супраневральную артерию, у Malocostraca – обычно субневральный сосуд. У хелицеровых и части многоножек брюшной сосуд соединен с передним концом сердца или с передней аортой одной парой дуг. У большинства Chilopoda он соединяется со спинным сосудом двумя парами дуг: передняя отходит от первой камеры сердца (в сегменте челюстных ножек), задняя – от последней камеры сердца. У сколопендры брюшной сосуд развивается в брюшном мезентерии, подобно брюшному и периневральному сосудам аннелид. Точная гомология его с каким-либо из этих сосудов трудно установима. Брюшной сосуд членистоногих входит в состав артериального отдела кровеносного аппарата и посылает артерии к различным органам, у многоножек – к конечностям. У Stomatopoda и Isopoda брюшной сосуд
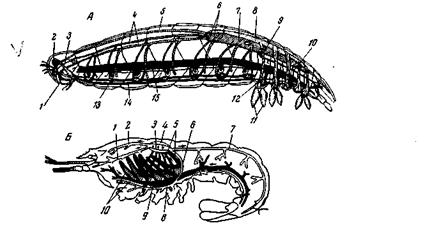
Рис. 158. Схемы кровеносного аппарата Malacostraca А – Isopoda: 1 – рот; 2 – мозг; 3 – окологлоточные коннективы; 4 – боковые артерии передних торакальных сегментов; 5 – аорта; 6 – боковые артерии шестого торакального сегмента; 7 – сердце; 8 – перикардиальный синус; 9 – бранхио-перикардиальные протоки; 10 – абдоминальные артерии; 11 – плеоподы; 12 – протоки, проводящие кровь от вентрального синуса к жаберным придаткам плеопод; 13 – брюшная нервная цепочка; 14 – субневральная артерия; 15 – вентральный синус; Б – Homarus (Decapoda): 1 – боковая головная артерия; 2 – аорта; 3 – сердце; 4 – перикардий; 5 – жаберные вены; 6 – arteria descendens; 7 – спинная артерия брюшка; 8 – arteria subneuralis; 9 – вентральный синус; 10 – жабры. Не изображена боковая внутренностная артерия (из Lang)
Наиболее примитивный тип хорошо развитой периферической системы артерий представляет формы с многочисленными гомономными метамерно-отходящими артериями. К этому типу больше всего приближаются Chilopoda с метамерными парами артерий, отходящими как от сердца, так и от брюшного сосуда на всем протяжении того и другого (рис. 161). Ветви головной аорты отходят также метамерно ко всем сегментам головы, включая, по-видимому, и тритоцеребральный. Более или менее гомономно отходят боковые артерии также у скорпионов и у Stomatopoda Гетерономная перестройка артериального отдела в той или иной мере свойственна также и всем паукообразным. Насекомые так же, как и низшие раки, отличаются значительной редукцией артериального отдела вообще. У насекомых артерии встречаются очень редко; примитивное метамерное расположение артерий, сравнимое с таковым высших раков или многоножек, описано для тараканов, имеющих две пары торакальных и четыре пары абдоминальных артерий (рис. 159); родственные им богомолы имеют четыре пары абдоминальных артерий; у некоторых других насекомых абдоминальные артерии заменены несколькими парами снабженных клапанами выходных остий. Обычно же аорта только в голове разделяется на две или несколько коротких веток. Изредка бывают артерии, направленные от сердца кзади (например, у поденок).
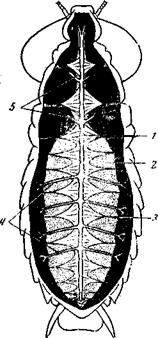
Рис. 159. Сердце и артериальная система Blaberus (Insecta, Blattoidea), вид с вентральной стороны 1 – сердце; 2 – дорсальная диафрагма; 3 – крыловидные мышцы; 4 – парные брюшные артерии; 5 – грудные артерии (по W.N. Nutting)
Венозный отдел кровеносного аппарата в более простых случаях совершенно отсутствует. Так, у насекомых сердце расположено в брюшке, где залегают также пищеварительный отдел кишечника и мальпигиевы сосуды. Засасывая через остии кровь, насыщенную пищевыми веществами и очищенную от экскретов, сердце прогоняет ее по аорте в голову, где расположены мозг и жевательная мускулатура. Под влиянием созданной таким образом разности давления кровь из головы стекает в грудь, где омывает локомоторную мускулатуру, и оттуда возвращается в брюшко. Строго дифференцированных путей для ее обратного оттока нет, но соединительная ткань образует ряд перепонок, так называемых диафрагм, которые до некоторой степени направляют потоки гемолимфы по более определенным путям. Наиболее развитой является брюшная, или супраневральная, диафрагма, идущая вдоль вентральной стороны брюшка; она обычно снабжена мышечными волокнами и в таком случае участвует в выполнении пропульсаторной функции, гоня кровь спереди назад. Сердце насекомых, как и спинной сосуд аннелид, гонит кровь, вообще говоря, сзади наперед. Однако уже со времен Мальпиги и Реомюра известно, что у некоторых личинок насекомых наблюдается периодическое извращение направления тока крови такое же явление описано и у взрослого комара (Anopheles maculipmnis), у которого сердце периодически меняет направление своей перистальтики, в течение известного времени посылая кровь из брюшка в голову, а затем в течение несколько более короткого периода засасывая кровь через аорту и», головы и изливая ее в брюшко. При более совершенном строении кровеносного аппарата, например у Decapoda (рис. 158, Б),кровь из артерий поступает в мелкие периферические синусы и оттуда постепенно собирается в большой вентральный синус. Отсюда кровь идет в жабры и из жабр по бранхиоперикардиальным каналам поступает в перикардий. Перикардий членистоногих, в отличие от такового моллюсков или хордовых, представляет собой кровеносную полость – перикардиальный синус, более или менее обособленный от остальной системы синусов. Сердце открывается в перикардий остиями. При наличии более или менее развитого венозного отдела перикардий совершенно теряет непосредственное сообщение с синусами тела и служит приемником для впадающих в него бранхиоперикардиальных или легочноперикардиальных сосудов. Такие сосуды, кроме высших раков, свойственны также мечехвостам и дышащим легкими паукообразным (скорпионы, телифоны, фрины, пауки). У этих паукообразных кровь из тканей собирается в большой вентральный синус, обмывающий легочные мешки. Здесь происходит насыщение крови кислородом, отсюда кровь по легочным венам поступает в перикардий и из него через остии – в сердце (рис. 160). Среди Tracheata зачатки венозного отдела кровеносного аппарата имеются только у Scutigeromorpha в виде лакун, подводящих кровь к перикардиальному синусу, в который погружены трахейные легкие. Слабое развитие кровеносного аппарата насекомых, несомненно, связано с развитием у них трахейного дыхания. Кровеносный аппарат крупных Malacostraca и части хелицеровых обспечивает не только снабжение тканей питательными веществами и освобождение их от экскретов, но также и снабжение их кислородом и освобождение от углекислоты. Кровь высших раков (Decapoda, Stomatopoda) содержит в растворенном виде дыхательный пигмент (гемоцианин). Ту же зависимость мы в известной мере видим и в других группах. Так, среди паукообразных снабженные трахеями группы (сенокосцы, лжескорпионы, Ricinulei, бихорки) имеют упрощенный кровеносный аппарат; сильнее всего он упрощен у бихорок, обладающих особенно совершенной трахейной системой. У них остается только сердце и передняя и задняя аорта, несмотря на относительно крупную величину бихорок. Точно так же и онихофоры, имеющие примитивную, но мощно развитую систему трахей, лишены каких бы то ни было сосудов, кроме сердца. У Scutigeromorpha с их централизованным дыхательным аппаратом (трахейные легкие) кровь, по-видимому, широко участвует в переносе кислорода и кровеносный аппарат (рис. 161) развит лучше, чем у других многоножек: сосуды толще, больше ветвятся, мускулатура сердца сильнее, сердце укорочено (назад тянется только до сегмента 13-х ног), лишено внутренних клапанов, т. е. начинает приобретать компактную форму. У насекомых за кровеносным аппаратом остается участие в несении трофической и экскреторной функций и обеспечение газообмена только небольших органов; газообмен всех более крупных и массивных органов обслуживается трахейной системой. Отсутствие централизованных дыхательных органов влечет за собой и отсутствие разветвленной системы артерий. Нет в крови у насекомых и дыхательных пигментов, если не считать некоторые специализованные случаи, вроде личинок Chironornus plumosus.Эти последние живут в илу водоемов, в условиях дефицита кислорода и имеют растворенный в гемолимфе гемоглобин, служащий здесь и для переноса кислорода по телу, и для образования запаса кислорода на периоды перерывов между дыхательными движениями, точь-в-точь, как у полихеты Nereis virens, no A. Lindroth: подобно Nereis, личинка Chironornus plumosus живет в U-образной трубке и качательными движениями создает в ней ток воды. Сходный образ жизни повел к далеко идущим конвергенциям в использовании дыхательного пигмента. Участие кровеносного аппарата насекомых в разносе кислорода по телу косвенно видно также и из наличия специальных приспособлений для насыщения гемолимфы кислородом. Таким приспособлением является, например, задняя камера сердца личинок комаров (сем. Gulicidae), сплошь оплетенная мелкими разветвлениями трахей, а также «кровяные жабры» многих личинок двукрылых.
Рис. 160. Схема строения Araneus diadematus (Araneina) 1 – рот; 2 – глотка; 3 – печеночные протоки; 4 – мальпигиевы сосуды; 5 – расширение задней кишки; 6 – задний проход; 7 – дилататоры глотки; 8 – слепые придатки кишечника; 9 – надглоточный ганглий; 10 – подглоточная нервная масса; 11 – сердце с тремя остиями; 12 – легкое; 13 – яичники; 14 – 16 – паутинные железы; 17 – паутинные бородавки; 18 – хелицеры; 19 – ядовитые железы; 20 – глаза; 21 – перекардий; 22 – легочные вены впадающие в перикардий; 23 – метамерные артерии; 24 – аорта (из Warburton)
Развитие системы трахей, утеря кровеносным аппаратом участия в газообмене крупных органов и связанное с этим его упрощение особенно характерны для насекомых. Эти черты принадлежат к основным организационным признакам этого класса. Система трахей в основном обеспечивает тканевое дыхание так же, как упрощенный кровеносный аппарат насекомых при малых размерах тела обеспечивает питание тканей и удаление экскретов. Но питание массивных органов при таком типе кровообращения вряд ли было бы возможно, с чем и связаны скромные размеры тела даже наиболее крупных насекомых. Самых крупных размеров среди членистоногих достигают как раз представители тех групп, которые обладают наиболее совершенно разветвленным кровеносным аппаратом: мечехвосты, эвриптериды – среди водных хелицеровых, скорпионы и многоножки – среди наземных членистоногих, десятиногие (например, Macrocheira из Anomura) – среди ракообразных. Некоторые эвриптериды имели 3 м в длину, Macrocheira достигает 3 м в размахе ног. Среди скорпионов Pandinus imperator достигает 20 см. Порядочные размеры имеют и некоторые многоножки: Scolopendra gigantea – 26,5 см, Scaphiostreptes (Diplopoda) – 28 см, Acantherpestes (карбон) – 50 см. Наряду с этим среди насекомых наиболее крупные жуки достигают 15 см, самые длинные палочники и стрекозы – 13 см, а громадное большинство насекомых отличается значительно меньшими размерами, измеряемыми миллиметрами. Теперь мы видим, что это обстоятельство не случайно: оно соответствует основным тенденциям развития насекомых и, в частности, связано с несовершенством их аппарата распределения.
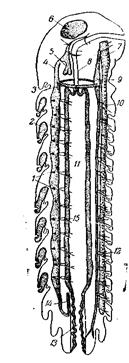
Рис.161. Кровеносный аппарат Scutiera (Chilopoda). Схема сагитального разреза животного 1 – остии сердца; 2 – веерные трахеи («трахейные легкие», свойственные Scutigeromorpha); 3 – дуга аорты; 4 – головная аорта; 5 – накачивающий аппарат; 6 – головной мозг; 7 – брюшная нервная цепочка; 8 – пищевод; 9 – брюшной сосуд; 10 и 12 – его разветвления; 11 – средняя кишка; 13 – задняя кишка; 14 – артерия задней кишки; 15 – боковые артерии сердца. Обратите также внимание на число тергитов, вдвое меньшее по сравнению с числом стернитов (из Догеля) Однако и среди членистоногих с хорошо развитым кровеносным аппаратом бросается в глаза колоссальная разница в максимальных размерах между наземными и водными формами: самые крупные эвриптериды имели в длину до 3 м, самый крупный скорпион – 20 см, другими словами, принимая во внимание более массивное телосложение эвриптерид, объем тела самых крупных из них в 4000 раз превосходит объем тела самого крупного скорпиона. Фактором, ограничивающим размеры тела всех наземных членистоногих, является мягкость покровов во время линьки: крупное тело при мягком наружном скелете должно было бы сильно деформироваться и деформировать своей тяжестью экзоскелет. Вопросы для самоконтроля
1. Примитивные формы распределительного аппарата. 2. Кровеносный аппарат немертин. 3. Кровеносная система аннелид. 4. Кровеносная система моллюсков. 5. Кровеносная система членистоногих.
ТЕМА 19
Поиск по сайту: |