|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Кровеносная система моллюсков
Моллюски резко отличаются от аннелид своей опорно-заполняющей тканью, имеющей в основном клеточный характер. Эта мезенхимная ткань всегда содержит большое количество щелей и лакун схизоцельного происхождения, заполненных кровью. Филогенетически за счет этих полостей и происходит развитие кровеносной системы моллюсков. Развитие это заключается в постепенной васкуляризации лакун (т.е. в постепенном превращении их в сосуды): первоначально неправильные полости постепенно получают характер синусов или каналов, по которым кровь легко может двигаться в определенном направлении; затем диаметр синуса уменьшается, а стенки его, образованные промежуточным веществом соединительной ткани, уплотняются и приобретают правильные контуры – синус превращается в кровеносный сосуд. В кровеносных сосудах некоторых моллюсков описывают и эндотелий. Стимулом для такого упорядочения лакунарной системы является возникающая в ней циркуляция жидкости. Самую простую форму кровеносного аппарата моллюсков мы находим у соленогастров (рис. 152, Б). Здесь сосудов нет. Имеются два продольных синуса – брюшной и спинной. Задняя часть спинного синуса впячена со спинной стороны в целом и снабжена мускулатурой, развивающейся за счет стенки целома. В силу этого она получает сократимость и гонит кровь сзади наперед вдоль спинного синуса. Этот сократимый участок спинного синуса получает название желудочка сердца.
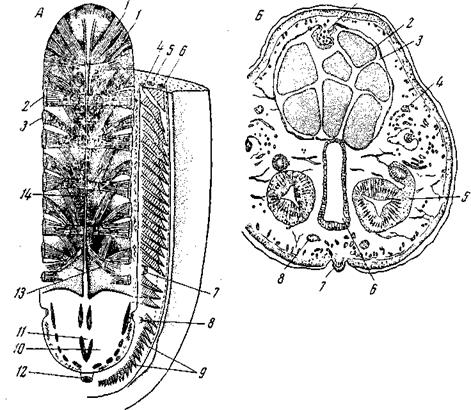
Рис. 152. Кровеносный аппарат Amphineura А– Acanthopleura echinata (Loricate): 1,2, 3– мышцы спинной стороны; обратить внимание на их метамерное расположение; 4 – приносящий жаберный сосуд; 5 – плевро-висцеральный нервный ствол; 6– уносящий жаберный сосуд; 7 – половое отверстие; 8 – мочевое отверстие; 9 – жабры; 10 – предсердие; 11 – желудочек сердца; 12 – задний проход; 13 – яйцеводы; 14 – аорта, отдающая метамерные артерии к мышцам и открывающаяся па своем переднем конце в головной синус; Б– схематический поперечный разрез через Stylomenia (Solenogastres): 1– сердце; 2 – половые продукты; 3– целом; 4– плевро-висцеральные нервные стволы; 5 – половые протоки;
Предсердии соленогастры не имеют, кровь в желудочек насасывается из выносящих жаберных синусов, еще лишенных правильных стенок. Сердце, как и у всех моллюсков, снаружи выстлано перитонеальным эпителием. Со стороны, обращенной к полости сердца, стенка этого последнего образована непосредственно мускулатурой. Такое строение стенки сердца свойственно и другим моллюскам. В остальном, строение кровеносного аппарата соленогастров чрезвычайно несовершенно и может обеспечить лишь весьма слабую степень циркуляции крови. Из хитонов Nuttalochiton hyadesi также, кроме сердца, не имеет никаких оформленных сосудов. У всех остальных хитонов спинной синус принимает характер правильного сосуда с хорошо выраженными стенками и получает название спинной аорты (рис. 152, А). Аорта идет прямо кпереди и широким отверстием изливается в головной синус – систему схизоцельных полостей, омывающих мозг. По пути аорта отдает ряд веток к половой железе и т.д. Головной синус отделен от остального тела диафрагмой, трубкообразное впячивание которой образует висцеральную артерию, доставляющую кровь к кишечнику и печени (у других моллюсков эти внутренности получают кровь непосредственно от аорты). Остальное кровообращение чисто лакунарное. Из мелких лакун кровь собирается в более крупные, главным образом в три продольных синуса ноги; отсюда она направляется в приносящие жаберные синусы, идущие вдоль внутреннего края мантийной полости. Пройдя через жабры, окисленная кровь собирается в выносящие жаберные синусы и по ним поступает в предсердия. Мускулистые предсердия перекачивают кровь в желудочек. Соединение толстостенного желудочка с тонкостенным предсердием широко распространено в животном царстве. Физиологический смысл этого приспособления заключается в следующем: для создания кровотока в артериальной системе необходим желудочек с достаточно толстыми мускулистыми стенками; растяжение его при диастоле требует значительного давления, которого не может быть в венозной системе. Предсердия обладают мускулатурой, достаточно сильной, чтобы растянуть расслабленные стенки желудка, и достаточно тонкой, чтобы быть способной растягиваться под влиянием венозного давления. Итак, мы видим у хитонов начало канализации системы кровеносных лакун, причем в обоих направлениях: с одной стороны, происходит упорядочение артериальной системы, несущей кровь от сердца, с другой – упорядочение венозной системы, приводящей кровь сначала к жабрам, а оттуда – к сердцу. Ближайшие к сердцу участки обеих систем упорядочиваются раньше, более отдаленные – позже. Среди брюхоногих мы еще встречаем формы со сравнительно короткими и слабо разветвленными артериальными и венозными стволами; головной синус лучше всего сохранился у наиболее примитивных Aspidobranchia – Fissurella, Haliotis. Наряду с этим высшие гастроподы имеют прекрасно разветвленную артериальную и венозную системы (рис. 153).
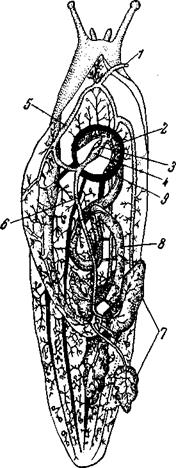
Рис. 153. Кровеносный аппарат Limax (Pulmonata Stylommatophora) 1 – половая артерия (обрезана); 2 – дыхательное отверстие; 3 – предсердие;
У большинства брюхоногих венозная кровь собирается в систему перивисцеральных синусов, окружающую кишечник, печень, и половую железу. Отсюда часть крови идет прямо в приносящий жаберный синус или сосуд, часть крови идет туда же, предварительно пройдя через почку (воротная система почек), некоторая часть крови попадает прямо в выносящий жаберный сосуд и через него – в предсердие. Таким образом, у большинства брюхоногих кровь в сердце преимущественно, но не чисто артериальная. Однако у некоторых Opisthobranchia, например у Gastropteron, вся венозная кровь, прежде чем попасть в сердце, проходит и через почку и через жабры. В этом они сходны с головоногими. Расположение приносящих и выносящих жаберных сосудов всецело зависит от устройства жаберного аппарата и в различных группах брюхоногих очень различно. У Pulmonata и других высших улиток движение крови, вызываемое втягиванием тела в раковину, гораздо более бурное, чем то, что может дать сокращение их сердца, но, с другой стороны, у этих животных перекачка крови из одного отдела в другой, производимая работой сердца, является важным составным элементом локомоторной деятельности Кровь улиток иногда содержит растворенные дыхательные пигменты: у Planorbis – гемоглобин, у Helix – гемоцианин. Кровь Loligo (Cephalopoda) тоже содержит гемоцианин. Среди головоногих у Nautilus появляются капилляры, но только в коже, и только здесь кровь переходит из артерий в вены, минуя лакуны. У Dibranchiata, в частности у Decapoda, система капилляров имеется не только в коже, но и в мускулатуре. Однако в области головы находится обширный синус, собирающий венозную кровь из головы и ноги. От этого синуса берет начало оформленная головная вена, которая подымается во внутренностный мешок и здесь делится на две полые вены, впадающие в жаберные сердца – сократимые мешочки, лежащие у основания жабр (рис. 154).
Рис. 154. Сердце и главные сосуды Spirula (Cephalopoda Dibranchiata) 1 – висцеральные нервы; 2 – соединение висцеральных нервов; 3 – рено-перикардиальное отверстие; 4 – почка; 5 – выносящий жаберный сосуд; 6 – приносящий жаберный сосуд; 7 – жаберное сердце; 8 – венозные придатки почки; 9 – половая артерия; 10 – мантийные артерии; 11 – левая брюшная вена; 12 – желудочек сердца;
Полые вены принимают в себя ряд венозных стволов от внутренностей. Проходя около почек, полые вены и другие венозные стволы вдаются туда гроздевидными расширениями – венозными придатками почек; они служат для очищения крови от экскретов. Жаберные сердца прогоняют венозную кровь через сосуды жабр, после чего она по жаберным венам попадает в предсердия и оттуда – в желудочек. Вся кровь, попадающая в желудочек, предварительно проходит через почки и через жабры. Octopoda отличаются от Decapoda лишь деталями. Таким образом, головоногие обладают весьма полным и совершенным кровообращением. Подобно рыбам, они имеют лишь один круг кровообращения, но тогда как у рыб сердце содержит венозную кровь, у головоногих оно содержит артериальную кровь. Затруднения, возникающие при необходимости прогонять кровь через две системы капилляров – капилляры тела и жаберные капилляры – преодолеваются у головоногих благодаря наличию дополнительных жаберных сердец. Совершенная система кровообращения и, в частности, весьма тонкое разветвление сосудов, доходящее до частичного перехода к замкнутому кровообращению, находится в связи с общим высоким уровнем организации головоногих и является одним из факторов, дающих некоторым из них возможность достигать гигантских размеров. Только при наличии системы капилляров возможно существование очень крупных животных, так как только при этих условиях обеспечено питание и дыхание массивных органов. Вообще, максимальные размеры тела, которых могут достигать представители какой-либо группы, больше всего зависят от степени совершенства распределительного аппарата. Чем совершеннее распределительный аппарат, тем крупнее могут быть обладающие им животные. Позвоночные с их высокосовершенным кровеносным аппаратом, с капиллярным кровообращением дают наиболее крупных животных, существующих на Земле. Головоногие и, в частности, Dibranchiata, обладающие частично капиллярным кровообращением и весьма совершенной циркуляцией крови, занимают второе место в лице таких гигантов, как Architeuthis. Как мы увидим дальше, это правило подтверждается и на примере различных групп членистоногих. Итак, организующим началом в кровеносной системе моллюсков является работа сердца. Сердце свойственно всем моллюскам, за исключением немногих, наиболее редуцированных форм – паразитические Pectinibranchia, Entoconchidae, карликовая деградированная голожаберная улиточка Rhodope). Сердце лежит в перикардии (целоме) у всех моллюсков, за исключением осьминогов и Anomia (Lamellibranchia Filibranchia). Перикардий обеспечивает свободу движения сердца и, в то же время, играет гидравлическую роль: при систоле желудочка объем жидкости внутри перикардия уменьшается, в силу чего создается отрицательное давление; это последнее вызывает приток крови из вен в предсердия, которые также лежат внутри перикардия (J. Ramsay, 1952). Сердце чаще всего лежит дорсально от задней кишки, изредка – вентрально от нее, а у Rhipidoglossa и у большинства пластинчатожаберных желудочек сердца пронизан задней кишкой, т.е. расположен периинтестинально. Этот последний тип строения является, вероятно, первичным. Можно полагать, что желудочек моллюсков гомологичен околокишечному синусу аннелид, последний описан и у Dinophilus, где он, в отсутствие постларвальных сегментов, помещается, как и сердце моллюсков, в ларвальном теле. Уменьшившись в размерах и приобретя роль мощного пропульсаторного органа, околокишечный синус стал желудочком сердца. Последний у большинства моллюсков эмансипировался от кишечника, сдвинувшись большей частью в дорсальном, реже – в вентральном направлении. Характерно, что среди форм, сохранивших периинтестинальное положение желудочка, находятся Rhipidoglossa В желудочек, как правило, симметрично открываются два предсердия. У Neopilina и Nautilus (рис. 155, Г)их четыре, у ряда брюхоногих – одно. Предсердия представляют мускулистые участки выносящих жаберных сосудов. У большинства моллюсков от сердца отходит одна передняя аорта (aorta cephalica); таковы боконервные (рис. 155, А),брюхоногие, Neopilina, Nautilus и немногие сравнительно примитивные пластинчатожа берные. У брюхоногих аорта дает мощную ветвь – внутренностную артерию (arteria visceralis). У двужаберных головоногих (рис. 155, В)и большинства пластинчатожаберных (рис. 155, Б)от сердца отходят две аорты – передняя и задняя. У некоторых головоногих прямо от желудочка отходят и некоторые другие артерии, обычно отходящие от аорты. Итак, в пределах типа моллюсков развитие аппарата распределения прослеживается от неправильной системы схизоцельных полостей, с едва начинающейся под влиянием зачаточного сердца циркуляцией, и до высокосовершенного кровеносного аппарата головоногих. Ни один из других типов беспозвоночных не дает среди своих современных представителей такого диапазона прогрессивных изменений кровеносного аппарата. Сравнивая архитектонику кровеносного аппарата моллюсков и аннелид, несмотря на все отличия, мы наблюдаем очень важные признаки сходства: исходным и первичным в обоих случаях является продольный, дорсомедиальный тракт, по которому кровь движется сзади наперед и который тесно связан в своем происхождении с околокишечным синусом. Различия в архитектонике кровеносных аппаратов обеих групп связаны с наличием у аннелид сильно развитого метамерного целома, между частями которого зажаты все сосуды и который определяет собой их расположение, тогда как сосуды моллюсков свободно развиваются путем упорядочения лакун в толще объемистой мезенхимы, заполняющей промежутки между органами.
Рис. 155. Схемы взаимоотношений между сердцем, жабрами и аортой у моллюсков А– Loricata; Б– Lamellibranchia; В – Cephalopoda Dibranchiata; Г– Cephalopoda Tetrabranchlata; Prosobranchia: Д– Rhipidoglossa (из числа Diotocardia Zygobranchia); E– Azygobranchia; Ж– Prosobranchia (из числа Monotocardia); З – Opisthobranchia Tectibranchia (no Hescheler)
Сходство в самых основных чертах архитектоники кровеносного аппарата подтверждает общее положение о близости исходных форм аннелид и моллюсков, а различия являются одним из совершенно очевидных проявлений далеко зашедшего расхождения обеих групп.
Поиск по сайту: |