|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Нервная система моллюсков
Нервный аппарат моллюсков в наиболее примитивной форме встречается у Amphineura (рис. 89). Мозг хитонов (рис. 89, А)представлен длинной полукольцевой комиссурой без дифференцированных ганглиев. Слабое развитие мозга хитонов несомненно стоит в связи со слабым развитием головных органов чувств, а слабое развитие этих последних – с малоподвижным образом жизни и травоядным питанием. От задних концов церебральной комиссуры отходят педальные и плевро-висцеральные стволы, соединенные между собой множеством поперечных комиссур. Первая из педальных комиссур проходит позади рта и носит название субцеребральной комиссуры. От нее отходит висцеральный отдел
Рис. 89. Центральный нервный аппарат Amphineura и Tryblidiida А – Acanthochilon discrepans (Loricata) (из Pelseneer); Б – Рrопeотeпiа sluiteri (Solenogastres);
передняя несет буккальные ганглии, иннервирующие спинную сторону ротовой полости и, может быть, кишечник; задняя несет субрадуляриые ганглии, иннервирующие вентральную сторону ротовой полости. Буккальный нервный аппарат моллюсков не гомологичен глоточному отделу нервного аппарата турбеллярий, поскольку рот и глотка турбеллярий не гомологичны таковым моллюсков и аннелид. Плевро-висцеральные стволы переходят друг в друга над анальным отверстием. Педальные стволы залегают у хитонов в толще мускулатуры ноги, которую они иннервируют, висцеральные – несколько дорсальней дна мантийной борозды. Таким образом, центральный отдел нервного аппарата уже у наиболее примитивных моллюсков значительно удалился от кожи. Нервный аппарат Neopilina galatheae (рис. 89) по уровню организации приближается к таковому хитонов, но менее примитивен. Число педальных комиссур низведено у Neopilina до двух (одна передняя и одна задняя), число комиссур между педальными и плевро-висцеральными стволами – до 10 пар. Имеются выраженные церебральные ганглии. Наряду с центральным нервным аппаратом, углубившимся под кожу, у всех моллюсков имеется и периферическое кожное сплетение (рис. 90), состоящее из всех типов нервных клеток и способное к самостоятельным рефлексам. Имеются и более глубокие периферические сплетения, например в аддукторах и других мускулах Lamellibranchia. Таким образом, у моллюсков произошла концентрация и уход в глубь тела не всех нервных элементов, а лишь части этих последних, которая и образовала собой центральный отдел нервного аппарата с отходящими от него периферическими нервами; другая часть осталась под кожей в виде диффузной сети, очень напоминающей первоначальную сеть кишечнополостных. В более примитивной форме аналогичный процесс мы видели у некоторых платод. Сложно развитое сплетение имеется и во внутренних органах моллюсков; с центральным отделом нервного аппарата оно стоит в связи через буккальные и висцеральные ганглии. Чрезвычайно существенно открытие чувствительных нервных клеток в составе эпителия кишечника Anodonta, позволяющее предполагать, что энтодерма моллюсков, подобно центральному фагоцитобласту ки- шечнополостных, образует нервные элементы. Что касается степени централизации нервного аппарата, мы могли бы сравнить хитонов с такими турбелляриями, как Mesostoma (Rhabdocoela), которая тоже имеет мозг и 4 продольных нервных ствола (хотя и негомологичные таковым хитонов). Вообще уровень развития нервного аппарата у хитонов и высших турбеллярий довольно близок, несмотря на коренное различие планов его строения в обеих группах. Таким образом, уровень организации двух аппаратов часто бывает возможно сравнивать даже в тех случаях, когда архитектонические планы строения обоих аппаратов в сравниваемых группах различны и самые части, составляющие эти аппараты, не гомологичны.
Рис. 90. Периферическое диффузное нервное сплетение в подошве ноги Helix pomatia (Gastropoda Pulmonata) (из Hanstrom)
В отношении тонкого строения центрального нервного аппарата аннелид, членистоногих, моллюсков и хордовых такое сравнение было в большом масштабе проведено А.А. Заварзиным и привело его к установлению «принципа параллелизма структур как одного из основных принципов морфологии»; следует только заметить, что параллелизм, так же, впрочем, как и дивергенция, свойственны эволюции не только структур, но всех вообще признаков организмов. Нервный аппарат соленогастров (рис. 89, Б, В)весьма близок к таковому хитонов, но представляет некоторые интересные отличия, частью уже описанные при рассмотрении архитектоники моллюсков. Здесь заметим лишь, что нервные клетки на протяжении стволов не располагаются у них равномерно, а концентрируются в местах отхождения комиссур, где образуют ганглии, а межлежащие участки стволов принимают характер простых проводящих путей, т.о. превращаются в коннективы. Однако у соленогастров этот процесс ганглионизации до конца не доведен, и на расположенных между ганглиями участках стволов некоторое количество нервных клеток у них все же остается. У большинства вышестоящих моллюсков все клетки центрального отдела нервного аппарата скопляются в компактные, резко очерченные ганглии, внутри которых образуется сплетение отростков (невропиль) и происходит контакт между отдельными невронами. Наоборот, промежуточные участки стволов, соединяющие два ганглия, полностью освобождаются от нервных клеток. Они состоят из параллельно идущих волокон, которые, лишь вступая в тот или иной ганглий, начинают ветвиться и контактируют с отростками других невронов. Ганглии представляют как бы телефонные станции, в которых возможно переключение, промежуточные участки стволов — как бы простые пучки проводов.
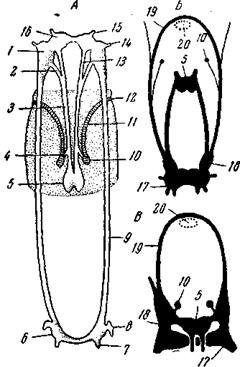
А – Haliotis (Rhipidoglossa), вид со спинной стороны, контуры тела нанесены пунктиром: 1 – глаза; 2 – ктенидии;
Ганглионизация является дальнейшим шагом по пути централизации нервного аппарата. Мы здесь опять видим укорочение внутрицентральных путей, если не в целом, то в пределах отдельных участков нервного аппарата. Нервный аппарат наиболее примитивных из Prosobranchia представляет известное сходство с таковым хитонов. Так, Haliotis (Aspidobranchia Rhipidoglossa) (рис. 91, А)имеет еще широкий и мало дифференцированный церебральный тяж, от которого идут нервы к глазам, головным щупальцам и волокна к статоцисту. У всех моллюсков эти органы иннервируются от церебрального отдела нервного аппарата, хотя волокна к статоцистам и идут обычно в составе церебро-педальных коннективов. Кроме того, от церебрального тяжа отходят и коннективы к буккальному отделу нервного аппарата. Вдоль ноги Haliotisтянутся два педальных ствола, соединенных метамерными комиссурами, первая из которых является субцеребральной комиссурой. В местах соединения субцеребральной комиссуры с педальными стволами отходят плевро-париетальные коннективы, перекрещивающиеся между собой и идущие к париетальным ганглиям. Эти последние лежат при основании ктенидиев, которые они иннервируют. От них идут плевро-висцеральные коннективы к висцеральному (абдоминальному) ганглию. Образованная этими тремя ганглиями дуга очевидно соответствует плевро-висцеральной дуге Amphineura. К месту ее соединения с педальными стволами подходят две пары коннективов, идущих от мозга; они очевидно представляют собой церебро-педальные и церебро-плевральные коннективы. Иногда возражают против гомологии плевро-висцеральной дуги высших моллюсков таковой Amphineura на том основании, что у первых дуга эта на своем заднем конце проходит под задней кишкой, тогда как у последних – над задней кишкой. Заметим, однако, что у Chaetoderma (Solenogastres) (рис. 89, В)имеется не только надкишечная комиссура, но и подкишечная, по существу представляющая последнюю из обыкновенных поперечных комиссур между стволами. Таким образом, разницу в положении комиссур в обеих группах моллюсков проще всего объяснить редукцией надкишечной комиссуры у Neopilina, у брюхоногих и других высших классов и сохранением подкишечной, т.е. последней из преанальных комиссур. Из поперечных комиссур, соединяющих у Amphineura педальные стволы с плевро-висцеральными, у Haliotis осталась только одна пара, а именно – на уровне субцеребралыюй комиссуры. У Haliotis эта плевро-педальная комиссура чрезвычайно укорочена, так что кажется, что педальные и плевро-висцеральные стволы непосредственно спаиваются между собой, но во многих случаях, например у Viviparus (Ar-chitaenioglossa), она очень наглядна. Итак, нервный аппарат Haliotis отличается от такового хитона хиастоневрией (черта вторичная), большим богатством церебральных органов чувств (отсутствие которых у хитонов может быть и вторично) и полной ганглионизацией плевро-висцеральной дуги, тогда как педальные стволы остаются почти на той же стадии развития, как и у хитона. В области плевро-висцеральной дуги обособились три ганглия: два париетальных и висцеральный. У большинства брюхоногих между париетальными и церебральными ганглиями бывают вставлены еще плевральные ганглии. У Haliotis они слиты с началом педальных стволов, а у других форм бывают соединены с ними парой плевро-педальных комиссур. Эти последние соответствуют одной из многочисленных комиссур, соединяющих педальные стволы Amphineura с плевро-висцеральными стволами. Резкое отличие наблюдается в строении периферических нервов, отходящих от плевро-висцеральных стволов, у Amphineura и брюхоногих. Прогрессивные изменения, наблюдаемые у вышестоящих переднежаберных, сводятся главным образом к дальнейшей ганглионизации нервного аппарата. Мозг Haliotis и большинства Aspidobranchia, как и мозг хитонов, слабо развит и представляет как бы поперечный участок ствола с расположенными на всем его протяжении нервными клетками; слабое развитие мозга связано с малой подвижностью низших аспидобранхий и слабым развитием головных органов чувств. Мозг большинства других улиток дифференцируется на два церебральные ганглия и комиссуру между ними (церебральная комиссура). Длинные педальные стволы свойственны лишь Aspidobranchia и отдельным представителям Pectinibranchia Architaenioglossa (Viviparus); строение педальных ганглиев Cypraea (Taenioglossa) лишь внешним образом напоминает педальные стволы низших улиток. У всех остальных улиток педальные стволы укорачиваются, конденсируются и превращаются в пару педальных ганглиев, соединенных между собой отныне единственной, педальной комиссурой, коннективами – с церебральными ганглиями и комиссурами – с плевральными ганглиями. Образование педальных ганглиев завершает ганглионизацию нервного аппарата брюхоногих и представляет существенный дальнейший шаг в процессе его централизации. Эпиподий, древнее наследие Rhipidoglossa, у вышестоящих форм больше не встречается. Наиболее примитивные из Opisthobranchia по организации своего нервного аппарата стоят еще на уровне переднежаберных (длинные коннективы, хиастоневрия). Среди Pulmonata некоторые следы хиастоневрии сохраняет Chilina и некоторые другие примитивные Basommatophora. В обоих подклассах, среди Opisthobranchia и среди Pulmonata, разыгрываются процессы раскручивания и укорочения коннективов, ведущие к ликвидации хиастоневрии. Однако наблюдаемое в обоих отрядах укорочение коннективов, ведущее к концентрации всех ганглиев вокруг пищевода, имеет и другое, более общее значение. Оно представляет дальнейший важный шаг в централизации нервного аппарата. У таких форм, как Tethys leporina (Nudibranchia),y которой все ганглии, кроме маленьких буккальных и висцерального, слились в одну общую массу, достигается одна из наиболее высоких степеней централизации нервного аппарата во всем животном мире (рис. 91, В). Однако надо помнить, что и у заднежаберных, как и у всех моллюсков, наряду с этим сохраняется и диффузное подкожное нервное сплетение.
Рис. 92. Схема диалинервного и зигонервного типов строения центрального нервного аппарата Prosobrancbia А – диалинервный тип; Б – правосторонняя зигоневрия; В – левосторонняя зигоневрия:
Нервный аппарат Lamellibranchia (рис. 93) построен по одному типу с таковым Gastropoda. Главные отличия заключаются, понятно, в отсутствии хиастоневрии, а кроме того, в упрощении, связанном со значительной редукцией головных органов чувств и упрощением ротового аппарата. По степени централизации своего нервного аппарата низшие пластинчатожаберные стоят приблизительно на уровне высших Prosobranchia: он полностью ганглионизован, но не сконцентрирован, коннективы длинные. Концентрация ганглиев наблюдается и здесь, а именно – в некоторых мономиарных семействах. Высшей степени она достигает у Lima. Однако, в отличие от гастропод, здесь все ганглии подтягиваются не к церебральным,
Scaphopoda по строению своего нервного аппарата приближаются к наиболее примитивным из Lamellibranchia, вроде Nucula (Protobranchia), но буккальный отдел развит лучше. Мы видели, что у брюхоногих централизация нервного аппарата сводится к двум процессам: ганглионизации стволов и укорочению коннективов и комиссур. По существу оба эти процесса между собой независимы, что видно при сравнении брюхоногих с головоногими. У брюхоногих сначала происходит ганглионизация стволов, а затем укорочение. У головоногих мы видим обратный порядок. Низшей ступени в развитии центрального отдела нервного аппарата моллюсков – длинных недифференцированных стволов – мы ни у кого из современных головоногих не находим. Зато укороченные нервные стволы, еще, однако, не дифференцированные на ганглии и коннективы, мы находим у Nautilus. Центральный отдел нервного аппарата Nautilus (рис. 94, А) состоит из трех дуг, образованных широкими тяжами, которые сплошь покрыты нервными клетками. Все тяжи укорочены и помещаются в голове. Один
Рис. 94. Центральный нервный аппарат головоногих А – Nautilus (Totrabranchia), вид сверху, схематизовано, щупальцевые нервы изображены частично: 1 – препедальное нервное кольцо; 2–- нервы пластинчатого органа; 3 – нервы к щупальцам; 4 – статоцист; 5 и 7 – нервы к передним и задним глазным щупальцам; 6 – глаза; 8 – абдоминальные ганглии; 9 – плевро-висцеральный тяж; 10 – церебральный тяж; 11 – буккальные ганглии; 12 – лабиальные ганглии; Б – Eledone moschata (Dibranchia Octopoda), вид с дорсальной стороны: 1 – лабиальные нервы; 2 – нервы рук; 3 – переднеглазничный нерв; 4 – зрительные ганглии; 5 – среднеглазничный нерв; 6 – верхние глазничные ганглии;
Кроме того, от церебрального тяжа отходит буккальная комиссура и от педального – препедальное кольцо. Последнее иннервирует часть ножных щупалец. Непосредственно от педальных тяжей иннервируются остальные щупальца и воронка, также представляющая одно из производных ноги, по мнению некоторых – гомолог эпиподия Rhipidoglossa. От плевро-висцерального тяжа, помимо многочисленных нервов к мантии и другим органам, отходят два коннектива к абдоминальным ганглиям, иннервирующим жабры и внутренности. Сдвиг педального кольца кпереди, наблюдаемый у всех головоногих, связан с характерным для них сдвигом кпереди и большей части зачатка самой ноги. В отличие от Nautilus, центральный отдел нервного аппарата всех двужаберных (рис. 94, Б)состоит не из недифференцированных тяжей, а из ганглиев. При этом коннективы и комиссуры чрезвычайно укорочены, так что основные ганглии – церебральные, педальные и висцеральные – сливаются вокруг пищевода в одну массу, внешне расчлененную лишь слабыми перетяжками. В то же время за счет периферического сплетения у головоногих дифференцируются дополнительные ганглии, отсутствующие у других моллюсков. Крупнейшими из них являются следующие: брахиальные ганглии, иннервирующие руки и стоящие в связи с педальными ганглиями, но также и непосредственно с церебральными; губные (лабиальные) или верхние буккальные ганглии, стоящие в связи с церебральными, но соединенные комиссурами также и с брахиальными; и, наконец, звездчатые ганглии (ganglia stellata), иннервирующие мантию и при помощи паллиальных нервов связанные с висцеральными ганглиями. Процесс ганглионизации периферического сплетения и вовлечение его в состав центрального нервного аппарата зашел у головоногих настолько далеко, что диффузные участки сплетения, если они и сохранились, не играют больше никакой роли ни в локомоторной деятельности животного, ни в игре хроматофоров кожи. Диффузное нервное сплетение сохранилось во внутренних органах (стенки кишечника и т.д.), а по некоторым данным, – и в той его части, которая связана с мускулатурой головы. В нервном аппарате головоногих имеются гигантские двигательные волокна. Они отличаются громадным диаметром (у Lоligo – 700 µ) и большой быстротой проведения, почти такой же, как в миэлиновых волокнах позвоночных. Гигантские волокна имеются у многих животных и обычно обслуживают особенно быстрые реакции бегства или укрытия в трубку, но у головоногих – нормальную функцию реактивного плавания. Впрочем филогенетически это последнее, вероятно, возникло из реакции, первоначально используемой при бегстве. Все головоногие – активные хищники; органы чувств и локомоторный аппарат двужаберных достигают высокого совершенства. В соответствии с этим для мозга головоногих характерно чрезвычайное развитие зрительных центров, громадный общий объем мозга, его сложное внутреннее расчленение и высокосовершенная микроархитектоника. Мозг головоногих защищен хрящевым черепом, у Nautilus – снизу, у двужаберных – почти со всех сторон. Мозг двужаберных и в особенности осьминогов – высшая точка в развитии центрального отдела нервного аппарата моллюсков и один из наиболее совершенных нервных приборов у всех беспозвоночных вообще. Вопросы для самоконтроля 1. Характеристика первичных нервных клеток. 2. Нервная система кишечнополостных. 3. Нервная система турбеллярий. 4. Эволюция нервной системы турбеллярий. 5. Строение нервной системы хитонов и моноплакофор. 6. Нервная система пластинчатожаберных и брюхоногих моллюсков. 7.Особенности строения и функционирования нервной системы головоногих моллюсков. ТЕМА 12
Поиск по сайту: |
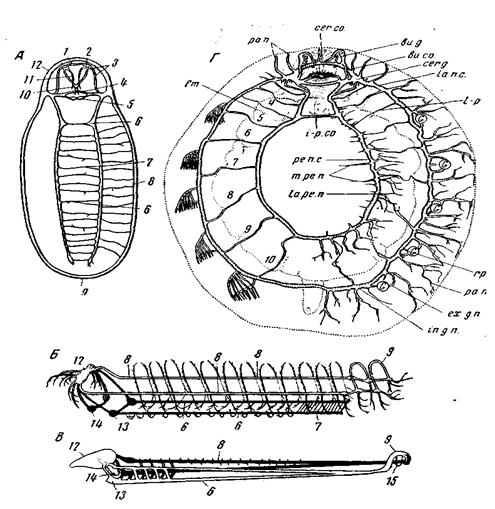


 Рис. 91. Нервный аппарат брюхоногих
Рис. 91. Нервный аппарат брюхоногих