|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Защитные приспособления кинобласта
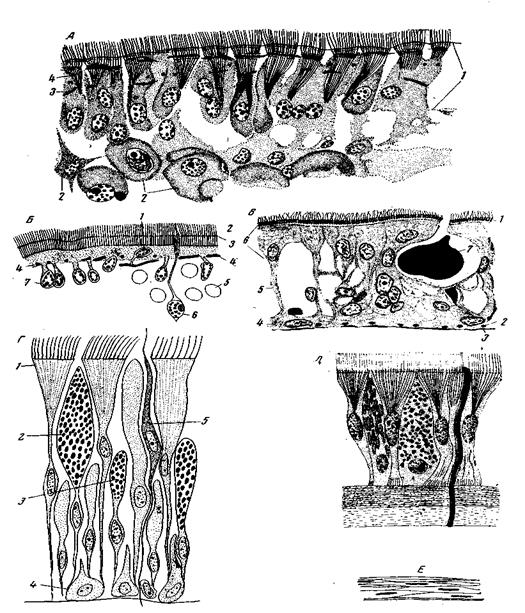
Образуя наружный слой тела животного, отграничивая его внутреннюю среду от наружной, кинобласт защищает организм от многообразных вредных воздействий внешней среды. Прежде всего он защищает внутреннюю среду от проникновения в нее различных вредных веществ и препятствует выходу из тела нужных этому последнему веществ. Далее, он защищает организм от проникновения различных живых и неживых частиц, от активного нападения врагов, от разрушения грубыми механическими воздействиями. Морфологически все эти функции выражаются, прежде всего, в правильном эпителиеобразном расположении клеток кинобласта у низших Меtazoa и клеток эпидермиса или наружного эпителия кожи (являющегося одним из производных кинобласта) – у вышестоящих форм. Дальнейшее морфологическое выражение защитная функция кинобласта находит в образовании желез, кутикул и пограничных пластинок. И оформленные секреты желез, и кутикула, и базальные пограничные пластинки в отдельных случаях могут, помимо защитной, приобретать также и опорную функцию, давая начало наружному скелету, а при дальнейшем осложнении организации – иногда и частям внутреннего скелета. Наконец, очень часто защитная функция эпидермиса усиливается благодаря интеграции его с нижележащими слоями, большей частью фагоцитобластического происхождения, образующими кутис (cutis) – соединительно-тканный слой кожи или в других случаях – кожно-мышечный мешок. Степень развития такого кожного покрова смешанного происхождения стоит в отрицательной корреляции с развитием кутикулы. Эпителиеобразное расположение клеток эпидермиса является общим правилом, это тот минимум функциональной структуры, который необходим для выполнения основной, отграничивающей роли наружного покрова. У части Acoela, обособленность эпидермиса от остальной паренхимы еще невелика. Его клетки не отделены от лежащей под ним паренхимы ни пограничной пластинкой, ни решеткой кожных мышц; пограничная пластинка у Acoela вообще отсутствует, мышечные волокна кожной мускулатуры у многих Acoela залегают интраэпителиально. Базальные концы эпителиальных клеток в некоторых случаях не образуют гладкой поверхности и вдаются между клетками паренхимы (рис. 75, А). Из этого примитивного типа легко выводятся четыре основных типа эпидермиса Bilateria. 1. Погруженный эпителий (рис. 75, Б). Апикальные части клеток слагаются в правильную эпителиальную пластинку, в которой иногда имеются, иногда отсутствуют клеточные границы. Наоборот, базальные концы клеток остаются независимыми друг от друга и включены в подлежащую ткань, не образуя обособленной от этой последней поверхности. Кожные мышцы остаются эпителиальными, располагаясь в эпителиальной пластинке, или обособляются от эпидермиса и ложатся на нижней поверхности эпителиальной пластинки. Этот тип эпителия свойствен многим турбелляриям – из отрядов Acoela, Alloeocoela и Triclada, всем паразитическим плоским червям и единичным формам из других групп. У всех этих форм, кроме бескишечных турбеллярий, эпителиальная пластинка бывает отграничена от нижележащих тканей базальной перепонкой. Погруженный эпителий паразитических платод отличается высокой степенью специализации. 2. Немертиновый эпителий, свойственный номертинам и в несколько отличной форме – части Acoela и других примитивных турбеллярий (рис. 75 В, Г). В этом случае эпителиальные клетки состоят из расширенных апикальных частей, слагающихся в сомкнутую эпителиальную пластинку, и узких, иногда ветвящихся базальных отростков, доходящих до пограничной пластинки или до решетки кожных мышц. Пространство между этими отростками заполнено железистыми клетками, клетками и волокнами опорного типа, нервными клетками и т.п. Таким образом, и здесь эпидермис слабо обособлен на базальной стороне от глубжележащих тканей, но здесь он входит в состав своеобразного кожного покрова, в образовании которого участвуют и элементы периферического фагоцитобласта. 3. Обособленный однослойный эпителий, свойственный большинству беспозвоночных (рис. 75, Д). В этом случае клетки эпидермиса принимают правильную призматическую форму, основания их подравниваются так же, как уже выравнены вершины. У Goelenterata и Acoela его клетки являются эпителиально-мышечными. У вышестоящих форм они дифференцируются на эпидермальные, остающиеся в эпителиальной связи между собой, и мышечные, утрачивающие связь с эпителием и располагающиеся под его базальной поверхностью. Среди кишечнополостных и бескишечных турбеллярий можно наблюдать промежуточные стадии этой дифференцировки. Наличие у Acoela всех трех первых типов эпидермиса, и постоянное отсутствие у них базальной перепонки показывает, что эта группа стоит очень недалеко от той ступени организации, на которой шло первоначальное обособление кинобласта и выработка основных типов эпидермиса. 4. Последний из основных типов эпидермиса представляет многослойный эпителий, свойственный позвоночным, а из беспозвоночных – одним Chaetognatha (рис. 75, Е).Этот тип эпидермиса представляет осложнение и усовершенствование предыдущего типа, «нормального» эпителия. Наружный эпителий бывает отграничен от нижележащих тканей базальной перепонкой (пограничной пластинкой из основного вещества). Нет ее только у Turbellaria Acoela и, у некоторых Macrostomida. Базальная перепонка, часто обладающая значительной прочностью, может стать основной опорой кожи. У многих турбеллярий и Monogenea за счет базальной перепонки эпителия половых протоков образуются шипы, стилеты и другие твердые части копулятивного аппарата, которые обычно ошибочно считают производными кутикулы. Существенную роль в защитной функции кинобласта играют кожные железы, слизистые и белковые, весьма многочисленные у кишечнополостных, турбеллярий, немертин, аннелид, моллюсков и многих других. Обычно это одноклеточные, бокаловидные или грушевидные железы, разбросанные по всей поверхности кожи или собранные в комплексы. Особенно постоянным у низших червей является головной комплекс желез, свойственный плоским червям, немертинам и многим личинкам аннелид. При погруженном эпителии все железистые клетки лежат в глубине паренхимы, и лишь протоки их пронизывают эпителиальную пластинку. При прочих типах эпидермиса часть железистых клеток входит в состав эпителия (дермальные железы), другие клетки погружаются под основную пластинку и лишь протоки их пронизывают эпителий (а д е н а л ь н ы е железы). Помимо защитной функции, кожные железы принимают на себя множество других: участие в локомоторной функции (например, железы подошвы наземных турбеллярий и брюхоногих улиток) и в прикреплении к субстрату (клейкие железы многих полипов и червей), участие в нападении на добычу (головные железы турбеллярий), выделение светящегося секрета (например у Chaetopterus из Spiomorpha). У турбеллярий часть слизистых желез вырабатывает секрет в виде оформленных зерен, шариков или палочек, известных под именем г и а л о-и д о в; в своей наиболее совершенной форме они называются р а б д и-т а м и (рис. 76, Б и В). Выбрасываемые из клеток на врага или добычу гиалоиды прилипают к нему, набухают, заклеивают и лишают свободы движения. За пределами турбеллярий рабдитные железы известны в эпителии хобота немертин. Еще более совершенный продукт выделения клеток эпидермиса представляют стрекательные капсулы книдарий (рис. 76, А).
Рис. 75. Основныетипы строения кожного эпителия А – эпителий, не обособленный от подлежащей ткани (паренхимы) – Oligocherus erythrophthalmus (Turbellaria Acoela): 1 – эпителиально-мышечные клетки; 2 – паренхимные клетки; 3 – мышечные волокна в эпителиально-мышечных клетках; 4 – базальный аппарат ресниц (но Беклемишеву) Б – погруженный эпителий Oligochoerus bakuensis; 1 – ядросодержащая часть мышечной клетки, сохранившая в виде исключения эпителиальное положение; 2 – ресницы; 3 – эпителиальная пластинка; 4 – волокна кожных мышц; 5 – контуры ядер паренхимных клеток; 6 – чувствительная клетка; 7 – погруженные в паренхиму ядра эпителия (ориг.); В – немертиновый эпителий Nemertoderma bathycola (Turbellaria Acoela): 1 – базальный аппарат ресниц; 2 – кольцевые мышцы кожи; 3 – продольные мышцы кожи; Особое морфологическое значение кожные железы приобретают в том случае, если выделяемый ими защитный секрет затвердевает и образует постоянные защитные образования. В некоторых случаях (многие полихеты и другие) эти защитные образования не соединены органически с телом животного и носят характер домиков. Мы можем проследить их эволюцию, начиная от простого применения секрета кожных желез для смазывания субстрата, по которому движется животное. Так, многие турбеллярии при ползании оставляют за собой слизистые нити, иногда турбеллярия может висеть на подобной нити, прикрепленной к какому-либо вышележащему предмету. Двигаясь в толще грунта, немертины, многие полихеты, наземные олигохеты выстилают слизью стенки прокладываемого хода и этим облегчают себе движение, уменьшая трение и защищая кожу от повреждения частицами грунта; фактически внутри хода создается слизистая трубка. У многих форм (например у Nereis virens) такой ход превращается в постоянное жилище, норку, выстланную слизью и несущую не только защитную функцию, но и гидрокинетическую, играя роль струенаправляющего приспособления. Вообще домики полихет представляют все переходы от временных и непрочных слизистых трубок к прочным и постоянным трубкам, образованным при участии склеенных вместе посторонних предметов (Onuphis conchilega из Eunicemorpha и другие) или из одного секрета, как, например, известковые трубки Serpulidae, достигающие большой прочности и у Spirorbiti – большой правильности форм. Трубки Terebellomorpha и Serpulimorpha образуются при участии секрета желез так называемых брюшных щитов торакса. Serpulidae, а также некоторые другие полихеты, никогда не покидают свою трубку и вынутые оттуда гибнут. Аналогичные ряды развития можно обнаружить и в других группах водных животных. У крыложаберных стенки домиков образованы затвердевшим секретом кожных желез головного щита и слагаются из характерных, веретенообразных отдельностей. Роgonophora имеют по всей длине туловища большое количество многоклеточных, колбообразных желез, глубоко вдающихся в целом, и, очевидно, служащих для выделения секрета, из которого строятся трубки этого животного. В других случаях животное остается прочно соединенным с выделенными защитными образованиями, которые превращаются, таким образом, в часть тела. Прежде всего таковы перисарк гидроидов, затем скелет Неxacorallia и, наконец, раковины моллюсков и плеченогих. Скелет Нехасогаlliа образован из углекислой извести (арагонита)
Рис. 76. Активно защитные приспособления эпидермиса А – стрекательные капсулы (глютинанты) гидры Pelmatohydra oligactis до и после выбрасывания нити (из Kukenthal); Б–В – расположение рабдитов в коже Trlclada (Б) и Polyclada (В): 1 – кольцевые мышцы кожи; 2 – продольные мышцы кожи; 3 – аденальные рабдиты; 4 – дорсовентральные мышцы; 5 – кожная железа; 6 – дермальные рабдитные клетки с рабдитами в протоплазме; 7 – базальная мембрана эпидермиса; 8 – аденальная железа (из Benham); Г – клептокниды; малая пенетранта гидры (Hydra vulgaris) в коже турбеллярии Microslomum lineare (Macrostomida):
Кольцевая складка подошвы, вдающаяся внизу внутрь тела полипа и концентрическая его наружной стенке, таким же образом выделяет теку. Аналогичным образом возникают и другие части, останавливаться на которых мы не будем. Существенно то, что весь этот скелет, топографически в значительной мере внутренний, морфологически является наружным: все его элементы выделяются исключительно наружной поверхностью эпидермиса, вдающегося внутрь тела разнообразными складками. Из всех моллюсков наиболее примитивным типом защитных приспособлений покровов обладают, по-видимому, Solenogastres. Вся дорсальная сторона их тела покрыта кутикулой; у некоторых видов она тонка и несет на поверхности обизвествленные спикулы, выделенные каждая одной эпидермальной клеткой. У других видов кутикула толста и в толще ее залегают многочисленные спикулы. Брюшная борозда и рудиментарная нога неомений во всех случаях лишены кутикулы и покрыты мерцательным эпителием. Loricata представляют следующую стадию в эволюции раковины: на определенных участках спинной стороны, дифференцировавшихся как сегменты раковинной железы, все клетки стали выделять известь и дают начало пластинкам раковины. К этому известковому слою перешла основная защитная функция, кутикула сохранилась на поверхности раковины в виде периострака. Периострак и tegmentum пронизаны органами чувств, эстетами, в чем можно видеть след происхождения раковины из совокупности отдельных спикул. Раковина состоит из 4 слоев: periostracum, tegmentum, articulamentum, hypostracum. Пластинки ее подвижно сочленены между собой. На периферии спинной стороны, в области perinotum, кутикула хитонов сохранила приблизительно то же строение, какое она имеет у соленогастров. У Conchifera раковина покрывает уже всю спинную сторону мантии и больше не пронизана эстетами; она состоит всего из трех слоев – periostracum, призматический и перламутровый слой и представляет единое монолитное образование. Наиболее примитивной известной нам формой раковины Conchifera являются ложкообразные или колпачкообразные раковины Tryblidiida. Метамерные мышечные отпечатки, нередко представленные восемью парами, может быть, указывают, что такая раковина возникла слиянием, восьми пластинок раковины примитивных Loricata. Раковина моллюсков несет защитную функцию, а также служит местом прикрепления мышц. У ряда брюхоногих из разных отрядов, в том числе у большинства заднежаберных, и у всех современных двужаберных головоногих раковина обрастает мантией и становится внутренней. При этом она теряет защитную функцию, но может сохранить опорную (многие Decapoda Dibranchiata) и гидростатическую функцию. В других случаях она становится рудиментарной и в конечном итоге полностью редуцируется (Octopoda, очень многие Gastropoda). В некоторых рядах брюхоногих раковина редуцируется и не проходя стадии внутренней раковины (киленогие – Taenioglossa Heteropoda и другие). Имеются случаи редукции раковины и среди хитонов. Мы видим многочисленные случаи исчезновения раковины или превращения ее во внутренний скелет в тех классах моллюсков, где основной ее функцией является защитная. Действительно, наряду с тенденцией к созданию раковин, панцирей и других приспособлений к пассивной защите, во всем животном царстве столь же распространена и противоположная тенденция к облегчению веса тела, нередко в связи с переходом к активной защите, связанной с усовершенствованием нервно-чувствительного Вторым типом защитных образований (наряду с секретами кожных желез) являются кутикулы. Кутикулой называют покрывающий поверхность эпителия защитный слой, представляющий не продукт выделения, а результат перерождения наружных слоев протоплазмы эпителиальных клеток. Впрочем, в ряде случаев отличить кутикулу от секрета трудно, и некоторые защитные образования возникают в результате совокупного действия обоих процессов: секреции и образования кутикулы. Несомненная кутикула впервые появляется у паразитических плоских червей и развивается у низших червей более или менее параллельно с редукцией мерцательного эпителия и ограничением его площади. Поэтому у турбеллярий она полностью отсутствует, у гастротрих и коловраток развита хорошо в тех местах тела, где отсутствует мерцательный эпителий. Кутикула нематод, несмотря на свою часто значительную толщину, гибка и остается нерасчлененной. Рост нематод и киноринх сопровождается во многих случаях сбрасыванием старой кутикулы, т.е. линькой, напоминающей линьку членистоногих. Есть ряд аналогий между кутикулой нематод и членистоногих в деталях: в строении чувствительных волосков и т.п. Тонкой и сравнительно просто устроенной кутикулой обладают аннелиды. Ни у нематод, ни у аннелид кутикула хитина не содержит. Большого развития достигает кутикула на поверхности цистидов мшанок, образуя наружный скелет этих последних; она содержит хитин, и у Stenolaemata и отчасти Gymnolaemata – обызвествлена. Однако высшую точку в смысле использования кутикулы в качество наружного скелета, несомненно, представляют членистоногие. Характернейшую особенность членистоногих представляет сплошная кутикула, дифференцированная на более толстые, твердые и негибкие склериты, являющиеся компонентами наружного скелета, и более тонкие и гибкие соединительные перепонки. Кутикула членистоногих состоит из хитина и протеина-артроподина. В склеритах этот последний уплотнен (склеротизован) благодаря дублению его о-хинонами, происходящему вскоре после линьки. У многих ракообразных, у Diplopoda и некоторых
Рис. 77. Схема разреза через кожу насекомого 1 – волоски; 2 – азальное кольцо волоска; 3 – эпикутикула; 4 – экзокутикула; 5 – псевдопоры; 6 – эндокутикула; 7 – эпидермис; 8 – основная перепонка эпидермиса;
личинок насекомых в кутикуле отлагаются значительные количества минеральных солей, в основном – углекислого кальция, но обызвествленная кутикула почти лишена артроподина. Тонкая структура кутикулы, в особенности в склеритах, вообще чрезвычайно сложна (рис. 77) и обеспечивает ей весьма совершенные механические свойства.
10.4. Дыхательные приспособления кинобласта
Громадное большинство животных обладает аэробным дыханием и, следовательно, нуждается в притоке кислорода из внешней среды к клеткам и тканям и в удалении углекислоты. У мелких организмов, даже и неподвижных, эти процессы могут совершаться при помощи одной диффузии. Гидра не нуждается в специальных дыхательных приспособлениях, так как почти все клетки ее тела находятся в непосредственном соприкосновении с окружающей водой, которая омывает не только эктодерму, но и энтодерму, проникая через рот в гастральную полость; здесь имеется лишь тканевое дыхание, и нет обособленных дыхательных органов. В таком же положении находятся и многие другие низшие многоклеточные – гидроиды, большинство сколецид, а также ряд карликовых форм из вышестоящих групп. С увеличением размеров тела одна диффузия обеспечить дыхание больше не может, и возникают те или иные механизмы, увеличивающие интенсивность газообмена выше тех пределов, которые может обеспечить одна диффузия. Все такие механизмы составляют дыхательные приспособления в широком смысле слова. Они делятся на три группы: 1) наружные дыхательные приспособления, усиливающие приток кислорода из внешней среды к дыхательной поверхности животного и удаление с этой поверхности углекислоты; 2) прямые дыхательные приспособления, облегчающие прохождение газов через пограничные эпителии животного; 3) внутренние дыхательные приспособления, обеспечивающие перенос кислорода от пограничного (дыхательного) эпителия к потребляющим тканям. Наружные дыхательные приспособления при помощи дыхательных движений создают потоки воды или воздуха, омывающие все тело животного или специализованные дыхательные эпителии. Прямые дыхательные приспособления выражаются, во-первых, в наличии специфической функциональной структуры дыхательных эпителиев, увеличивающей их проницаемость для кислорода и углекислоты, и, во-вторых, в увеличении поверхности видоизмененных таким образом эпителиев. Прямые дыхательные приспособления входят, как правило, в состав настоящих дыхательных органов и рассматриваются ниже. У губок и кишечнополостных животных встречаются только наружные дыхательные приспособления, и прежде всего в виде гидрокинетических органов. У менее подвижных водных животных наружные дыхательные приспособления встречаются как правило. Большинство таких животных создает вокруг себя токи воды. Создаются они темп же способами, которые служат и для передвижения животных: движением ресниц, мышечными движениями всего тела (например, Nereis, Tubifex, личинки Chironomidae) или его придатков (например, многие ракообразные). Очень сильно развиты и разнообразны мерцательно-гидрокинетические приспособления у иглокожих, среди Eleutherozoa – у звезд и ежей. В состав вододвижущего аппарата, кроме вододвижущих органов, могут входить и струенаправляющие приспособления, как мы видели на примере двустворчатых и асцидий. В большинстве случаев создаваемые животным токи воды имеют для него, как мы уже видели, разностороннее значение: они приносят кислород, пищу (у большинства фильтраторов и седиментаторов) или запахи пищи (некоторые полихеты). Однако в ряде случаев их функция чисто дыхательная (например, у Tubificidae, являющихся исключительными илоедами). Широкое распространение настоящие дыхательные органы получают только у полихет. Жабры полихет бывают двух основных типов: кровяные и целомические. И те, и другие представляют торчащие в воду, большей частью разветвленные, выросты стенки тела. Наиболее типичные кровяные жабры – гребенчатые параподиальные жабры Eunicidae (рис. 78, А). У сидячих полихет они встречаются чаще, чем у бродячих. Все кровяные жабры характеризуются богатым разветвлением кровеносных капилляров. Наоборот, в коже целомических жабр кровеносных капилляров не бывает. Они свойственны тем формам, у которых роль внутренней дыхательной среды играет не кровь, а полостная жидкость, как, например, Nephthydidae, Glyceridae, Capitellidae и т.п. Многие полихеты, в особенности блуждающие, вообще лишены жабр (Nereidae, Phyllodocidae и многие другие), причем это отсутствие в одних случаях является следствием редукции, а в других быть может и первичным. У Nereis функцию дыхательных органов выполняют параподии, а также обособленные участки эпителия спинной и брюшной стороны сегментов тела, пронизанные густой сетью кровеносных капилляров. У олигохет и пиявок параподиальные и головные жабры, свойственные полихетам, отсутствуют, но у ряда отдельных форм из обоих классов возникли вторичные жабры (рис. 78, Б, В),также представляющие выросты стенки тела и иногда снабженные весьма совершенной системой приносящих и выносящих сосудов (Branchellion из Rhynchobdellea). Из пиявок жабры имеются у многих Ichthyobdelidae (Rhynchobdellea), из олигохет – у отдельных форм из разных семейств (Dero и другие). Первичные жабры моллюсков, или ктенидии (рис. 79) гребенчатой формой отчасти напоминают параподиальные жабры Eunicidae, однако имеют по 2 ряда лепестков. Подобно жабрам полихет, ктенидии хитонов также расположены метамерно по бокам туловища, и предположение о гомологии их жабрам полихет очень соблазнительно. Neopilina имеет 5 пар метамерных жабр, несущих по одному ряду хорошо развитых лепестков. Ктенидии трех первых пар имеют, кроме того, по второму ряду лепестков, однако рудиментарных. У других моллюсков ктенидиев бывает одна пара, кроме Nautilus, имеющего 2 пары. Подвергаются редукции ктенидии у большинства соленогастров, у ряда специализованных групп улиток и у части Lamellibranchiа (отряд Septibranchia) (рис. 80, Г). Своеобразную эволюцию проделывают ктенидии у большинства пластинчатожаберных и у некоторых Ргоsobranchia. Из пластинчатожаберных только у Protobranchia ктенпдии сохраняют свою обычную форму (рис. 79, Г; рис. 80, Л). В других отрядах этого класса они разрастаются во всю длину мантийной полости, и лепестки их вытягиваются в длинные, сложенные пополам нити (отряд Filibranchia, рис. 80, Б). При дальнейшем развитии эти нити сливаются между собой и образуют пластинчатые, сетчатые жабры, характерные для наиболее многочисленного отряда Eulamellibranchia (рис. 80, В). Впрочем, как уже упомянуто выше, основной функцией жабр высших двустворчатых является вододвигательная и фильтровальная, и самое превращение жабр из ктенидиальных в пластинчатые очевидно обусловлено колоссальным возрастанием их вододвигательной функции, вызвано переходом от сбора донного детрита к питанию при помощи фильтрации.
Рис. 78. Жабры аннелид A – кровяные параподиальные жабры Eunice aphroditoides (Polychaeta); Б – анальные жабры у Dero incisa (Oligochaeta, Naididae); В – боковые жабры у Ozobranchus jantseanus (Hirudinea Ichthyobdellidae) (по Ока); 1 – жабры Рис. 79. Ктенидии моллюсков, полусхематически (по Ray-Lankester) А – Chiton; Б – Sepia; В – Fissurella; Г – Nucula; 1 – приносящий жаберный сосуд; 2 – продольный мускул жабры; 3 – парные жаберные листочки; 4 – выносящий жаберный сосуд
Рис. 80. Жабры Bivalvia Схематические поперечные разрезы тела в области жабр. А – Protobranchia; Б – Filibranchia; В – Eulamellibrancuia; Г – Septibranchia (по Hescheler); Д – переход дыхательной функции к мантии; сильное развитие кровеносных сосудов мантии молодой устрицы (Ostrea lurida)l1 – дополнительное сердце (сократимый участок мантийного сосуда); 2 – радиальные сосуды; 3 – мускул-аддуктор раковины; 4 – сердце; 5 – внутренности; 6 – отводной канал мантийных сосудов; 7 – очертания жабр, видимых сквозь мантию; 8 – почка; 9 – краевой кровеносный синус мантии (по Hopkins)
В ряде случаев происходит даже ослабление дыхательной функции жабр, что видно из обеднения их кровеносными сосудами; в таких случаях дыхательная функция частично переходит к мантии, кровоснабжение которой соответствующим образом усиливается и видоизменяется (например, у устриц Ostrea lurida, рис. 80, Д). Таким образом, эволюция жабр LamellibrancMa характеризуется частичной или полной сменой функции и в последнем случае субституцией их в качестве дыхательных органов стенками мантийной полости. Предположение о гомологии между жабрами членистоногих и параподиальными жабрами полихет очень соблазнительно. Особенно правдоподобным оно представляется, если принимать экзоподит и эндоподит раздвоенной конечности ракообразных за гомологи спинной и брюшной ветви параподии полихет, как это делали многие, большей частью старые авторы. Само по себе предположение о гомологии ног членистоногих параподиям полихет правдоподобно и подтверждается сходством наружной мускулатуры примитивных конечностей членистоногих и параподии полихет. Но некоторые авторы, как, например, R. Snodgrass (1938), отрицают даже и эту гомологию; и хотя точка зрения Snodgrass является крайностью и мало обоснована, вопрос о гомологии жабр обеих групп остается пока открытым. Жабрами обладают личинки многих живущих в воде насекомых. Для всех рассмотренных типов жабр первичноротых характерно то обстоятельство, что они представляют выросты стенки тела, свободно омываемые водой и большей частью гребенчатые или разветвленные. Другими словами, для них характерно максимальное развитие поверхности соприкосновения с наружной дыхательной средой. Ряд наблюдений над личинками комаров показывает, что при обеднении окружающей среды кислородом и других затруднениях дыхания происходит компенсаторное увеличение жабр, а следовательно и дыхательной поверхности. У мелких личинок комаров относительная величина жабр меньше, чем у крупных. Отсюда понятно, что мелкие животные, с их более крупной удельной поверхностью, очень часто довольствуются одним кожным дыханием, обходясь без того увеличения дыхательной поверхности, которое связано с наличием обособленных дыхательных органов (Copepoda среди раков, ряд клещей среди паукообразных и т.п.).
Рис. 81. Жабры личинок насекомых А – личинка Taeniopteryx nebulosa (Plecoptera) с коксальными жабрами (по Lauterborn);
10.5. Защитные и дыхательные приспособления кинобласта при
Воздух содержит во много раз больше кислорода, чем вода, даже если вода находится в равновесии с атмосферой. Скорость диффузии кислорода в воде во много тысяч раз меньше, чем в воздухе. Таким образом, воздух во много раз лучший источник кислорода, чем вода. Поэтому очень многие водные животные не могут обойтись без наружных дыхательных (гидрокинетических) приспособлений, без которых наземные животные тех же размеров и того же уровня организации большей частью обходятся. «Не легко переменить дыхательную среду, но обеспечение 21 % кислорода является для такой перемены мощным стимулом». Переход животных к наземному образу жизни совершился, вероятно, различными путями. По мнению G. Colosi, общей причиной появления высших групп наземных животных является интенсификация обмена веществ, идущая в ногу с повышением организации. Усиление обмена вызывает повышенную потребность в кислороде, и данная группа животных получает стремление все больше вылезать из воды и постепенно переходить к наземному образу жизни. По мнению Колози, на этой стадии эволюции находятся в настоящее время, например, очень многие крабы. Однако непосредственный переход из моря прямо на сушу, если и возможен, то главным образом для высших животных, позвоночных и высших раков, обладающих достаточной осмотической самостоятельностью внутренней среды, при которой атмосферные осадки не угрожают животному гибелью от действия гипотонической жидкости. Примитивные представители легочных моллюсков, в частности Siphonariidae, также являются морскими, литоральными формами, и их выходу на сушу может быть помогла та защита, которую обеспечивает им раковина. В целом, низшие Metazoa легче переходят на сушу из пресных вод, нежели прямо из моря. Основная масса наземных турбеллярий, нематод и аннелид связана с пресноводными, а не с морскими представителями тех же групп. И большой дорогой при переходе их из воды на сушу явилось, как правильно утверждает М.С. Гиляров, обитание в почве, в лесной подстилке, во мху – во в лажных местообитаниях, представляющих ряд промежуточных ступеней между условиями водоема, с одной стороны, и открытых наземных местообитаний – с другой. Постепенное завоевание какой-либо группой животных этих промежуточных ступеней вело ее шаг за шагом от водного образа жизни к наземному и от организации, приспособленной к жизни в воде, к организации настоящего наземного животного. Очень многие группы животных так и не пошли дальше влажных, укрытых местообитаний и являются, в сущности, полуназемными формами. При переходе к наземному образу жизни перед животным возникает необходимость в ряде приспособлений и, в частности, возникает задача защиты от испарения, которая распадается на две частные задачи: сохранение запаса воды в теле и защита от высыхания дыхательных поверхностей. Ряд наземных животных является «несовершенно» приспособленным к наземной жизни: они не защищены от испарения и потому могут жить только в среде, воздух которой насыщен водяными парами. Сюда относятся все наземные турбеллярии, немертины и голые моллюски, лишенные всякой кутикулы, защищенные только кожными железами и живущие лишь в постоянно влажной среде. Сюда же относятся «наземные» коловратки, нематоды и тардиграды, снабженные тонкой кутикулой, которая их не защищает от высыхания, и активные лишь в периоды высокой влажности, но способные большей частью переносить высыхание в состоянии анабиоза. Немногим лучше защищены наземные олигохеты и пиявки. Снабженные раковиной моллюски большей частью активны тоже лишь во влажном воздухе, но некоторые из них могут населять биотопы с резкими колебаниями влажности, проводя сухие периоды в пассивном состоянии внутри раковины (например, Xerophila из Pulmonata). «Совершенно» приспособленные к наземной жизни беспозвоночные встречаются только среди членистоногих, кутикула которых при наличии некоторых приспособлений может создать вполне достаточную защиту от испарения. Onychophora, морфологически стоящие между членистоногими и аннелидами, по степени защищенности от высыхания также занимают промежуточное положение: в одинаковых условиях Peripatus теряет вдвое меньше воды, чем дождевой червь, и вдвое больше, чем сколопендра такой же величины. У насекомых, помимо толщины кутикулы, основным приспособлением против высыхания является наличие эпикутикулы, образованной тонким слоем воскоподобного вещества, непроницаемого для паров воды. Вторая задача – защита от высыхания дыхательных поверхностей – связана с тем, что газообмен, по-видимому, возможен только через влажную поверхность. У всех животных дыхательный эпителий всегда бывает влажен. Сухих дыхательных органов не бывает. Colosi обозначает это положение как принцип постоянства дыхательной среды и дает ему следующую, с виду несколько парадоксальную, формулировку: все животные, независимо от их сред обитания, всегда дышат в воде. Под этим он разумеет, что молекулярно тонкий слой воды, покрывающий влажную поверхность дыхательного эпителия наземных животных, является посредником между этим последним и атмосферой. В силу принципа постоянства дыхательной среды, с переходом к дыханию в сухой среде наружные, или эвагинированные дыхательные органы, становятся невозможными. «Несовершенно» приспособленные наземные животные, живущие хотя и в воздухе, но насыщенном влагой, могут еще дышать всей поверхностью тела. В то же время богатство атмосферного воздуха кислородом позволяет на суше обходиться без обособленных дыхательных органов и наружных дыхательных приспособлений даже животным, относящимся к таким группам, водные представители которых нередко обладают и тем, и другим (аннелиды). Однако при переходе к жизни в открытых местообитаниях и к дыханию сухим воздухом становится необходимым образование внутренних, или и н в а г и н и р о в а н н ы х, дыхательных органов, эпителий которых был бы защищен от высыхания. И такого типа дыхательные органы действительно возникают у всех «совершенно» приспособленных наземных животных, тогда как у водных животных типичные инвагинированные дыхательные органы встречаются сравнительно редко (хордовые, личинки стрекоз, офиуры, Blastoidea и т.д.); однако очень часто эвагинированные жабры водных животных бывают защищены складками, предохраняющими их от загрязнения илом и в то же время играющими роль струенаправляющих приспособлений, усиливающих обмен воды около жабр (Mollusca, Crustacea и др.). Из моллюсков настоящими наземными животными являются только легочные улитки (Pulmonata). Они выработали себе полый дыхательный орган путем редукции ктенидия, перехода дыхательной функции ко всему эпителию мантийной полости и уменьшению ее наружного отверстия до степени небольшого дыхальца. Мантийная полость превращена в легкое. Ее стенка покрыта густыми разветвлениями кровеносных сосудов, кровь которых и участвует в газообмене. Среди ракообразных сравнительно немногие живут на суше, и те большей частью имеют лишь примитивные приспособления к воздушному дыханию. Десятиногие, особенно крабы, представляют хороший пример использования при переходе на сушу целого ряда приспособлений, выработанных еще в воде. Ноги их еще в воде были приспособлены для бегания по дну. Жабры защищены от механических повреждений и засорения спускающимися с боков складками панциря и помещаются, таким образом, в особых жаберных камерах; при выходе из воды эти складки защищают их и от высыхания; толстый панцирь защищает от высыхания тело. Благодаря этому многие крабы перешли к полуназемному, а некоторые к наземному образу жизни (Ocypoda среди Brachiura; Birgus среди Anomura). В крайних случаях происходит рудиментация и даже исчезновение жабр, и дыхательная функция переходит к стенкам жаберной полости, превращенной, таким образом, в легкое (Birgus). У наземных Amphipoda и Isopoda анатомически выраженные приспособления к воздушному дыханию также большей частью сводятся к изменению формы жаберных ножек и созданию большей защиты для жабр. У части мокриц (Oniscoidea) расширенные в виде крышечки экзоподиты передних плеопод снабжены впячиваниями в виде тонких, кустообразных разветвленных трубочек, сходных с трахеями наземных членистоногих и образующих трахейные легкие. Их дыхательная функция (поглощение кислорода) экспериментально доказана. Наземные хелицеровые, объединяемые под названием паукообразных, представляют два типа дыхательных органов: легкие и трахеи. Природа легочных мешков паукообразных была выяснена Ray Lankester, который доказал их гомологию жаберным ножкам водных Chelicerata. До него легочные мешки считались видоизменением трахей. Легкие паукообразных (рис. 82, В, Г)представляют собой мешки, открывающиеся поперечными щелями на брюшной поверхности мезосомы; полость их заполнена большим количеством тонких листков, отходящих от передней стенки легкого и немного не доходящих до его задней стенки. Эти листки у скорпионов и пауков весьма напоминают жаберные листочки Limulus, а у телифонов – скорее таковые трилобитов. При эмбриональном развитии легких паукообразных сначала в соответствующих местах образуются в виде бугорков зачатки жаберных ножек. Затем позади каждого бугорка образуется ямка – зачаток полости легкого, а на задней поверхности бугорка появляются складки – зачатки первых жаберных листков (рис. 82, Л, В). Дальше конечность не растет в высоту, а ямка при ее основании углубляется. Весь передний край этой ямки соответствует задней поверхности ножки (состоящей почти из одного преэпиподита) и в соответствии с этим покрывается складками, дающими начало листкам легкого. Однако эмбриологи нередко отказывают этим листкам в точной гомологии с жаберными листками Limulus на основании того курьезного довода, что они не сами врастают в полость легкого, а, наоборот, полость вдается в толщу ножки складками, разделяющими лепестки, как будто это различие имеет какое-либо значение. Итак, легкие паукообразных и по способу своего развития, и по строению, как и по положению, представляют видоизменение жаберных конечностей водных хелицеровых, приросших краями к покровам и углубившихся своей задней поверхностью в глубь тела. Это далеко идущее приспособление к защите дыхательного эпителия от высыхания, хотя никаких новообразований в составе легкого нет, а происходит лишь изменение уже существующего органа. Наряду с этим у паукообразных чрезвычайно резко проявляется самая, может быть, своеобразная черта кутикулярного покрова – его реакция на переход к наземному образу жизни путем образования трахей. Мы встречаемся с независимым образованием трахей в самых различных отрядах паукообразных, в разных местах тела и за счет различных источников. Среди акариморфных клещей примитивные, слабо склеротизованные формы, живущие в постоянно влажной среде (лесная подстилка и пр.), лишены трахей; у вышестоящих форм, вместе со склеротизацией кожи, возникают трахеи, и притом – у различных групп различным образом. То обстоятельство, что трахеи не являются чем-то унаследованным от общих предков наземных членистоногих, как думали когда-то, а независимо возникали в отдельных группах, уже давно стало ясно на примере паутинных пауков (Araneina). У примитивных пауков (Liphistiomorpha, Mygalomorpha) и в некоторых семействах Araneomorpha (например, у Pholcidae) трахей вовсе нет. У большинства двулегочных (Dipneumones s. Araneomorpha) имеются две пары трахейных стволов на месте задней пары легких (рис. 82, Д).Однако в некоторых семействах пауков (Caponiidae, Telemidae и другие) на месте передней пары легких также образуются трахеи; все относящиеся сюда семейства мало родственны между собой и не образуют никакой естественной группы, так что утрата передней пары легких и замена их трахеями происходила в различных группах двулегочных пауков неоднократно и независимо. Как показывает история развития, из задних двух пар трахейных стволов Dipneumones латеральная пара развивается за счет зачатков легких, а медиальная представляет видоизменение простых мышечных апофиз. Мышечные апофизы – отростки кутикулы, отходящие внутрь тела и служащие для прикрепления мышц. Становясь полыми и тонкостенными и начиная ветвиться, апофизы одной пары превращаются в трахеи. У других паукообразных встречаются случаи образования трахей за счет межсегментных перепонок. Таким образом, источники образования трахей весьма различны. Из всех этих данных вытекает следующий важный вывод: очевидно невозможно говорить о монофилетическом происхождении трахей в разных классах членистоногих, поскольку у близких форм и даже у одного и того же животного они нередко возникают несколькими независимыми путями.
Рис. 82. Дыхательные органы паукообразных А и Б – две стадии развития легких Lycosa (Araneina) (из Lang); В – схема строения легкого паука (Araneina) (по Mac Leod); Г – разрез через свободные края трех легочных листочков Euscorpius flavicaudis (Scorpiones) (из Kastner); Д – трахеи крестовика Araneus diadematus (Araneina) (из Lang); E – трахейный аппарат Galeodes arcmeoides (Solifugae), животное вскрыто со спинной стороны (по Kastner); 1 – полость легкого; 2 – гиподерма легочных мешков; 3 – входы из преддверия легкого в пространство между листочками; 4 – кровеносные лакуны внутри листочков; 5 – хитиновые ворсинки, препятствующие слипанию листочков, у края последних сплетенные в «кружево»
Кроме паукообразных, типичные трахеи имеются у многоножек и насекомых, весьма примитивные трахеи имеются у Onychophora, зачаточное образование трахей мы видели у мокриц. Одним словом, всякий раз, когда какая-либо группа покрытых кутикулой животных переходит к воздушному дыханию, у нее появляется тенденция к возникновению трахей. У Onychophora (рис. 83, А)трахеи представлены пучками тончайших неветвящихся трубочек (1–3 µ в диаметре), которые открываются наружу общим отверстием (стигмой) и глубоко вдаются внутрь тела. Стигмы в неисчислимом множестве покрывают всю поверхность тела, включая и голову, но многочисленней всего на спине. Расположены они беспорядочно, хотя иногда намечается некоторое расположение продольными рядами. У насекомых и многоножек в прототипе трахеи образуют также кустики, но значительно более мощных, чем у Onychophora, стволиков, обычно ветвящихся. Они всегда расположены строго метамерно, по одной паре в каждом сегменте. Дыхальца лежат дорсальней места прикрепления конечностей. Трахейная система насекомых отличается возрастающей гетерономностью и интеграцией. В прототипе по одной паре дыхалец имеется в каждом сегменте, начиная с лабиального и кончая девятым брюшным. Однако в лабиальном сегменте стигмы известны лишь у зародыша Apis и у взрослых Sminthurus (Collembola). Зародыш Leptinotarsa (Coleoptera) имеет три грудных и девять брюшных пар стигм, взрослые Heterojapix (Campodeoidea) – три грудных и восемь брюшных, Lepismatidae и многие взрослые Pterygota – две грудных и восемь брюшных, Machilidae – две грудных и семь брюшных (рис. 83, В). Дальнейшая олигомеризация встречается уже и у Apterygota (три пары торакальных стигм у Сатpodea, две пары – у Eosentomon, одна пара головных – у Sminthuridae) и составляет правило у Holometabola. В прототипе отдельные кустики трахей, или трахеомеры, независимы между собою. Такое строение полностью сохранилось у Campodea и Eosentomon и почти полностью – у Machilidae. Между тем, образование анастомоз между трахеомерами в виде исключения встречается уже у некоторых Diplopoda и, как правило, – у Chilopoda Epimorpha. У Japigidae все трахеомеры каждой стороны соединены между собой продольными стволами, но поперечная комиссура имеется всего одна. У Lepismatidae имеются такие же продольные стволы и по одной поперечной комиссуре в каждом сегменте. Pterygota по строению трахейной системы в главном сходны с Lepismatidae. У многих из них происходит дальнейшее сокращение числа дыхалец, и в крайнем случае их остается всего одна пара, например у личинок комаров (Culicidae), сохраняющих, однако, и несколько пар рудиментарных дыхалец. Тонкие трахеолы, в отличие от больших трахейных стволов, не анастомозируют никогда. У многих летающих насекомых (Diptera, рис. 83, Г, Hymenoptera) главные стволы образуют большие воздухоносные мешки или же в мелких стволиках возникает множество мелких пузырьков, также служащих для уменьшения удельного веса тела (Coleoptera). Чрезвычайной сложности и разнообразия достигают у насекомых и некоторых паукообразных дыхальца или стигмы в связи с образованием различных защитных и, замыкательных механизмов. С физиологической точки зрения трахеи представляют дыхательный аппарат весьма своеобразного типа. При высокой степени развития трахейной системы тонкие трахейные стволики и трахеоли бесчисленными разветвлениями непосредственно оплетают все внутренние органы, проникают внутрь некоторых клеток и непосредственно обеспечивают газообмен тканей. Гемолимфа в значительной мере теряет роль дыхательной среды.
Рис. 83. Трахеи Onychophora и Arthropoda А – разрез через стигму и пучок трахей Peripatopsis capensis: 1 – кутикула; 2 – эпидермис; 3 – трахея (из Lang); Б – трахеи передней части тела Julus mediterraneus (Diplopoda); пучки трахей отходят от трахейных карманов (4), развитых, начиная с IV сегмента; во II и III сегментах нлеразвитые трахейные карманы (2 и 3) (по Krug); В – Machilis (Thysanura) – примитивный тип трахейного аппарата насекомых (из Lang); Г – Musca domestica (Diptera) – высокодифференцированный трахейный аппарат высших насекомых; вид сбоку для показа главных воздухоносных стволов и воздушных мешков: 1 – летательные мышцы; 2 – передняя грудная стигма (из Berlese);
Если легкие скорпиона или почти неветвящиеся трахеи паука представляют централизованный дыхательный аппарат, функция которого состоит в насыщении гемолимфы кислородом и освобождении ее от углекислоты, то трахеи насекомого или бихорки представляют децентрализованный дыхательный аппарат, пронизывающий все тело, всюду проникающий и обеспечивающий тканевое дыхание без посредства гемолимфы. Эта особенность трахейной системы оказывает большое влияние на всю организацию насекомых и, по всей вероятности, на основные тенденции эволюции этого класса. Обмен воздуха в дыхательных органах многоножек, большинства паукообразных и многих насекомых всецело совершается путем диффузии или, самое большее, при участии сжатия и расправления трахей при изгибаниях тела животного. Однако многие насекомые, а из паукообразных бихорки и Ricinulei, совершают настоящие дыхательные движения. Небольшие дыхательные движения совершают и пауки. У жуков происходит ритмическое опускание и подымание мягкой дорсальной поверхности брюшка, у стрекоз – такие же колебания их узких стернитов, у кузнечиков тергиты брюшка приближаются к стернитам и удаляются от них и т.д. При каждом повышении внутрибрюшного давления воздушные мешки или толстые, мягкостенные стволы трахей спадаются, при понижении давления вновь заполняются воздухом. Этим путем обеспечивается смена воздуха в главных трахейных стволах; в разветвлениях она и здесь совершается путем диффузии. Смена воздуха в главных трахейных стволах увеличивает приток кислорода к дыхательному эпителию трахеол, и потому скелетно-нервно-мышечные механизмы, обеспечивающие дыхательные движения, следует признать наружными дыхательными приспособлениями. Возникновение их явно связано с повышением интенсивности обмена веществ у высших наземных членистоногих. Кроме этих последних, дыхательными движениями из всех наземных животных обладают только позвоночные и некоторые легочные улитки.
Вопросы для самоконтроля
1. Основные функции кинобластов. 2. Мерцательно-локомоторные и вододвижущие приспособления губок и гребневиков. 3. Мерцательно-локомоторные и вододвижущие приспособления кишечнополостных и низших червей. 4. Вододвижущие аппараты полихет и моллюсков. 5. Характеристика типов мерцательно-вододвижущих аппаратов. 6. Характеристика типов эпидермиса. 7. Кожные железы, их функции. 8. Защитные образования эпидермиса (раковина, скелет, кутикула). 9. Пути перехода животных к наземному образу жизни. 10. Морфология дыхательных органов наземных беспозвоночных. ТЕМА 11
СТРОЕНИЕ НЕРВНОЙ СИСТЕМЫ КИШЕЧНОПОЛОСТНЫХ, НИЗШИХ ЧЕРВЕЙ И МОЛЛЮСКОВ
11.1. Нервная система кишечнополостных
Компоненты нервной системы кишечнополостных принадлежат к трем типам: это чувствительные клетки, ассоциативные и двигательные. Первые сохраняют свое эпителиальное положение, остальные его утратили и располагаются субэпителиально, на одном уровне с мышечными отростками миоэпителия. Все они образуют многочисленные отростки, вступающие в контакт с другими нервными клетками (невронами), а отростки двигательных клеток, кроме того, и с мышечными клетками. В отличие от невронов вышестоящих форм, нервные клетки кишечнополостных, по меньшей мере их ассоциативные клетки, не поляризованы: все их отростки равноценны и не разделяются на неврит и дендриты. Все нервные клетки соединены между собой в единое сплетение, охватывающее все тело животного и представляющее его нервный аппарат. У гидроидных полипов это сплетение имеет диффузный характер и образует лишь не особенно резко выраженные сгущения вокруг рта, а у гидр – также и на подошве. Мы уже упоминали о замечательной особенности кишечнополостных, заключающейся в том, что у них нервное сплетение развито не только за счет кинобласта, но и за счет фагоцитобласта – эпителия кишечника. Оба сплетения, эктодермальное и энтодермальное, переходят друг в друга в области кишечного рта. Первичной формой нервных клеток являются лежащие в эпителии чувствительные клетки, из которых выводятся как ассоциативные, так и двигательные клетки. Естественней всего представить себе прототип нервной системы кишечнополостных (рис. 84, A) в виде сети расположенных в эпителии клеток, снабженных на вершине воспринимающими приспособлениями, а при основании – отростками, при помощи которых каждая из этих клеток вступает в контакт как с себе подобными, так и с мышечными клетками. Непосредственная связь чувствительной клетки с мышечными представляла бы осуществление простейшей мыслимой дуги рефлекса – одночленной. Такого типа связи действительно описываются в щупальцах актиний. Связь всех клеток нервного сплетения между собой обеспечивает реакции на чисто местные раздражения со стороны более обширных областей тела. Теперь представим себе (рис. 84, Б), что часть этих универсальных клеток утратит непосредственную связь с мышечными клетками и сохранит эпителиальное положение и связь с другими нервными клетками; что другая их часть утратит непосредственную связь и с поверхностью
Рис. 84. Схема возникновения чувствительных, ассоциативных и двигательных нервных клеток кишечнополостных (Б) из универсальных нервных клеток прототипа (А) 1 – чувствительные окончания; 2 – синапсы в местах контакта различных невронов;
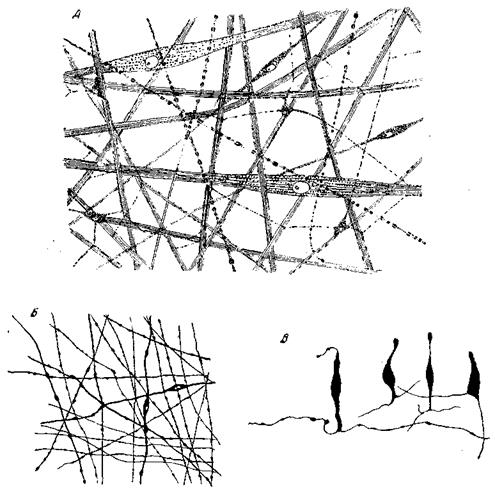
Диффузный нервный аппарат кишечнополостных (рис. 85) представляет весьма примитивный тип нервного аппарата. В нем отсутствует разделение на центральный отдел и периферический, отсутствует специализация среди ассоциативных клеток, отсутствуют длинные, состоящие из одних отростков проводящие пути. Сеть проводит раздражение по всем направлениям, причем эта передача раздражений совершается от нейрона к нейрону. В то же время каждый из них связан и с двигательными клетками. В силу этого распространяющаяся по нервной сети из какой-либо точки волна возбуждения на всем своем пути сопровождается и волной мышечного сокращения. Полип, как правило, неспособен реагировать на возбуждение, возникшее в какой-либо точке его тела, сокращением удаленных от нее мышц, если раньше не прореагируют все мышцы, расположенные между обеими точками.
Рис. 85. Нервный аппарат Cnidaria Нервное сплетение медузы Rhizostoma (Scyphozoa): А – эктодсрмальное; Б – энтодермальное; на рис. А изображены также и мышечные волокна (по Bozler); В – группа энтодермалъных чувствительных клеток актинии Bunodes gemmaceus (пo Grosel)
Передача раздражения в нервной сети совершается несравненно быстрее, чем при отсутствии нервных элементов у губок (у актиний – со скоростью 4–15 см в 1 секунду); но несравненно медленней, чем проведение в периферических нервах высших Metazoa, где не приходится преодолевать вставленных по пути синапсов, т.е. мест контакта между отростками разных невронов. Однако и у актиний существуют быстро проводящие пути, обеспечивающие, например, реакцию защитного сокращения животного. В них скорость проведения возбуждения несколько больше 1 м в секунду. Морфологически эти пути представлены кольцом на оральном диске и продольными тяжами в мезентериях (см. рис. 86).
Вероятно синапсы на этих путях существуют, но переход возбуждения через них перманентно облегчен (J. Ramsay, 1952).
Вторая особенность, вытекающая из строения диффузного нервного аппарата, заключается в том, что любой кусочек, вырезанный из тела полипа, способен к самостоятельным рефлексам, способен давать мышечные сокращения в ответ на раздражение, в отличие, например, от отрезанной ноги насекомого. Это происходит от того, что при диффузном, абсолютно нецентрализованном нервном аппарате полипов каждый малей- ший кусочек тела всегда содержит весь нервный механизм, необходимый для осуще- ствления рефлекса – чувствительную, ассо- циативную и двигательную клетки и мускул-исполнитель, связанные между собою.
Поиск по сайту: |