|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Приспособления кинобласта
В хорошем соответствии с теорией происхождения Metazoa от Zooflagellata стоит тот факт, что мерцательная функция кинобласта осуществляется при помощи жгутов прежде всего в наиболее примитивных группах многоклеточных, у губок и многих книдарий, а также у личинок книдарий, иглокожих и Acrania. Из взрослых Bilateria исключительно жгутиковыми мерцательными эпителиями обладают, например, плеченогие, широко распространены жгутиковые эпителии у взрослых иглокожих, в эндостиле асцидий. У большинства Bilateria мерцательную функцию несут ресницы, являющиеся результатом полимеризации и упрощения жгутов. Из групп Metazoa, обладающих ресничками, Turbellaria Acoela являются одной из примитивнейших, и в соответствии с этим базальные аппараты их ресниц сложнее и больше напоминают базальные аппараты жгутов, чем это свойственно большинству Metazoa; у Acoela и Polyclada мы иногда встречаем такую флагеллятную черту, как прикрепление ризопластов к ядру (рис.73, Д). В ряде случаев все тело многоклеточного животного бывает сплошь покрыто мерцательным эпителием, как мы видим у жгутиковых бластул различных животных, у планул кишечнополостных, у взрослых турбелля-рий и немертин и т.д. В других случаях мерцательный эпителий сосредотачивается на определенных участках тела и образует сложно дифференцированный мерцательный аппарат. Как использование, так и типы этого аппарата в различных группах животных очень различны. Губки, по примитивности многих черт своей организации стоящие близ самого корня Metazoa, в отношении использования кинобласта значительно отличаются от остальных многоклеточных. Все личинки губок плавают при помощи жгутов кинобласта, выстилающего все или почти все тело личинки (у паренхимул) или образующего собой анимальную половину личинки (у амфибластулы Calcarea). В противоположность личинке взрослая губка является сидячим животным. Однако, хотя она прикреплена к субстрату и передвигаться не может, жгутики продолжают работать в течение всей ее активной жизни. Если губка не двигается, движение жгутиков заставляет двигаться воду. Таким образом мерцательно-локомоторная функция переходит в мерцательно-гидрокинетическую. Кинобласт из органа передвижения превращается в орган создания тока воды и привлечения вместе с ней пищи. Приспособление к сидячему образу жизни ведет у губок к самой глубокой ломке организации, какую мы только видим у кого-либо из сидячих животных. Более мощное привлечение пищи внутрь организма достигается у губок тем, что кинобласт уходит внутрь тела, образуя там жгутиковые камеры. Благодаря этому у них происходит извращение зародышевых листков; кинобласт оказывается внутри толщи фагоцитобласта; фагоцитобласт дифференцирует на своей поверхности слой эпителиеобразно расположенных пинакоцитов, который служит наружным покровом, но негомологичен таковому остальных Metazoa, а представляет образование Если считать губок, как это обычно делается, первой нисходящей ветвью, отделившейся от главного ствола Metazoa, наличие у них извращения зародышевых пластов чрезвычайно поучительно. Оно представляется при этом допущении ярким примером того правила, что чем ниже организация какой-либо группы животных, тем более глубокие изменения вызывает в ней всякое приспособление. Разрешая сравнительно простую задачу, создавая приспособление для привлечения воды, губки буквально вывернулись наизнанку. Более сложные животные (иглокожие, аннелиды, мшанки, усоногие и др.) приспособляются к выполнению той же задачи с гораздо меньшими изменениями своей организации. Одним из существенных признаков прогрессивной эволюции является возрастающая эмансипация организма от прямого и непосредственного воздействия отдельных факторов среды, что мы наблюдаем и в области сравнительно-физиологической, и в области эволюционно-морфологической. Гребневики являются животными сравнительно крупных размеров. Между тем плавать при помощи ресничек могут обычно только очень маленькие животные, ибо реснички – слабые органы, и для того, чтобы они могли поддержать тело животного в воде, должно существовать определенное соотношение между весом этого тела и числом ресничек, т.е. между массой животного и величиной его поверхности. Гребневики – глубоко консервативная группа. Они сохранили чисто мерцательный способ передвижения, почти не переходя к плаванию при помощи мускулатуры, и лишь немногие из них перешли к сидячему образу жизни или ползанию. Каким образом гребневики поддерживают в воде свое тело при помощи мерцательного движения? Это достигается прежде всего усовершенствованием самих ресниц. Вместо отдельных ресниц гребневики имеют гораздо более эффективные гребные органы – гребные пластинки, образованные каждая слиянием двух рядов ресниц. Затем у гребневиков происходит уменьшение удельного веса, что достигается, во-первых, высоким процентным содержанием воды в их теле (благодаря чему оно сильно теряет в механической прочности), и, во-вторых, быть может, регуляцией ионного состава тканевой жидкости в сторону снижения концентрации тяжелых ионов, например SO4.. Гребные пластинки образуются не на всем теле гребневиков, а располагаются восемью меридиональными рядами. Сохраняющиеся у них простые реснички также располагаются лишь в определенных частях тела. Таким образом, у гребневиков происходит не только интеграция самих ресничек в пластинки, но и дифференцировка их расположения, – явление, широко распространенное у всех вышестоящих форм. Эта дифференцировка представляет один из простейших примеров той концентрации отдельных функций кинобласта в пределах отдельных его участков, о которой мы выше упоминали, как об одном из основных путей эволюции первичных пластов Metazoa. Большинство книдарий ведет сидячий образ жизни, и жгуты или реснички кинобласта, когда сохраняются, не играют у них больше локомоторной роли, а, как и жгуты у губок, служат для приведения в движение воды. Вообще говоря, вызываемые жгутами или ресницами токи воды могут выполнять у различных животных три главные задачи: во-первых, обновляя воду около поверхности тела, они облегчают дыхание и удаление экскретов; во-вторых, они очищают поверхность тела от оседающих из воды на нее частиц; во многих случаях эта функция имеет жизненное значение, например у видов Madreporaria, живущих в тихой воде лагун; и в-третьих, те же токи могут привлекать ко рту пищевые частицы. Кишечнополостные первично являются хищниками-хватателями и ловят сравнительно крупную добычу при помощи щупалец и подвижных краев рта. Таким способом питаются все Hydrozoa и большинство представителей других классов, в том числе все наиболее примитивные их представители. Однако ряд кишечнополостных вторично перешел к питанию более или менее мелкорассеянной пищей, приносимой ко рту ресничками. Из сцифомедуз это доказано для взрослых Aurelia и ризостомид, из гребневиков доказано для Мпеmiopsis и весьма вероятно в отношении некоторых других Lobifera, а также и Cestoidea. Среди актиний, наряду с хищными формами, имеющими хорошо развитые щупальца и слабо развитый ресничный покров, вроде Urticina, есть ряд микрофагов со сравнительно слабо развитыми щупальцами и хорошо развитым мерцательным покровом, приносящим ко рту пищевые частицы с поверхности диска и щупалец. Плавают при помощи ресниц только мелкие турбеллярии, не свыше 3 мм в длину. Более крупные могут еще продвигаться при их помощи по субстрату, но чем крупнее турбеллярия, тем меньшую роль в ее передвижении играют ресницы, тем большую – мускулатура. У турбеллярий, плавающих при помощи ресничек, движение этих последних зависит от нервной системы и может приостанавливаться или изменять направление под влиянием нервных импульсов. У всех крупных турбеллярий реснички несут гидрокинетическую функцию (смена воды вокруг тела). В этом случае они постоянно бьют в одном направлении спереди назад и не поддаются больше регулирующему влиянию нервной системы. Их движение является автономным. Из паразитических платод Temnocephala несут реснички только на щупальцах, взрослые сосальщики и ленточные черви совершенно лишены ресничного покрова. Однако личинки сосальщиков (мирацидии Digenea, многие личинки Monogeinea) и ленточных червей (ликофоры и корацидии примитивных семейств) бывают покрыты мерцательным эпителием и плавают при его помощи. Из других сколецид движение при помощи ресниц сохраняют отличающиеся своими ничтожными размерами коловратки и Gastrotricha. Однако вместо сплошного мерцательного покрова и у тех, и у других сохраняется, как и у некоторых турбеллярий, лишь полоска мерцательного эпителия на брюшной стороне. У коловраток этот первоначальный тип расположения ресниц, свойственный донным нотомматидам, у многих коловраток он служит не только для плавания, но на долю его выпадает и новая задача – привлечение пищи ко рту. Большинство полихет сохраняет более или менее развитый мерцательный аппарат и во взрослом виде, однако для плавания он служит лишь у упомянутых карликовых форм; основная его функция – гидрокинетическая. Как строение мерцательного аппарата, так и назначение его в различных семействах полихет – очень различны. В ряде случаев он образован отдельными группами или полосками ресничных клеток на сегментах, в других случаях имеется сплошной ресничный эпителий на головной лопасти или на тех или иных придатках. Ряды ресничных клеток, сидящие на сегментах тела и параподиях афродитид, служат для создания токов воды к защищенным элитрами жабрам; дыхательное значение реснички имеют и у ряда других форм. У Serpulimorpha имеется мерцательная бороздка, идущая от ануса к передней части тела и служащая для удаления экскрементов из жилой трубки. Реснички, сидящие на головных придатках большинства сидячих форм, служат для подгона ко рту пищевых частиц, но в тех случаях, когда частицы приносятся сильным током воды, последняя используется и для дыхания, и для удаления отбросов. Мерцательные аппараты, служащие для обмена воды около поверхности тела, широко распространены и у моллюсков. Особенное значение они приобретают в тех случаях, когда обеспечивают циркуляцию воды в мантийной полости, например у большинства Lamellibranchia и Gastropoda Prosobranchia; в этих случаях нередко возникают специальные струенаправляющие приспособления, как например образованные краем мантии сифоны, эпиподиальная борозда аспидобранхий (Risbec, 1955) и др. У соленогастров ресничным эпителием покрыт рудимент ноги, расположенный в виде валика вдоль брюшной борозды Neomenioidea, у хитонов и брюхоногих – подошва ноги, жабры и мантийная полость, у брюхоногих – сверх того нередко и другие места тела; голожаберные (Nudibranchia) обычно покрыты ресничным эпителием сплошь. Пластинчатожаберные также почти сплошь покрыты ресничным эпителием, за исключением наружной поверхности мантии, служащей раковинной железой, эпителия сифонов и т.д. У головоногих мерцательный эпителий сильно редуцирован, и обновление воды вокруг жабр достигается сокращениями мускулатуры мантии. Во многих из перечисленных случаев работа ресниц имеет главным образом значение усиления обмена воды близ поверхности тела, что существенно прежде всего для дыхания животного. Наряду с этим, как мы уже видели, у многих животных работа ресниц используется и для подгона пищевых частиц ко рту или фильтрующему аппарату. Эта функция для мерцательного аппарата Enterozoa является вторичной. Под ее влиянием он часто приобретает большую сложность и совершенство. Существуют 5 главных типов мерцательно-вододвижущих аппаратов, служащих для привлечения пищи и независимо возникавших в различных группах билатерий. 1. Реснички расположены по всей поверхности тела или на поверхности значительных его областей и гонят воду, слизь и приставшие к ней частички по направлению ко рту (Lobifera, многие актинии) или ко вторичным ротовым отверстиям, образовавшимся при зарастании рта (взрослая Aurelia). Вероятно, такого же типа гидрокинетическим аппаратом обладали и Amphoroidea (рис. 73). 2. Реснички располагаются вокруг рта, обычно на венце щупалец, свободно торчащем в воду, и в своей совокупности создают ток воды, направленный ко рту. При этом щупальца могут быть расположены простым кругом, как у сабеллид, у мшанок Staenolaemata и Gymnolaemata, или образуют более сложный лофофор, как у Phoronis, Phylactolaemata и некоторых других; иногда щупальца перисто разветвлены, как у большинства Serpulimorpha. Аппараты этого типа конвергентно возникают в различных группах: у коловраток (в наиболее выраженной форме у Stephanoceros), у Podaxonia, Kamptozoa и у Polychaeta, причем у последних неоднократно. Так, мерцательный ловчий аппарат Serpulimorpha образован разветвлениями головных придатков (пальп), тогда как аналогично функционирующий ловчий аппарат Hermellidae (Spiomorpha) образован гребневидными рядами «ротовых нитей», сидящих на дорсолатеральных лопастях, или воротничке, образованном разрастанием четырех первых сегментов торакса (рис. 73). 3. Реснички расположены в приводящих бороздах, которые при работе обращены к субстрату; движение ресниц гонит пищевые частицы вдоль борозд ко рту; питание – не за счет толщи воды, а за счет поверхности грунта. Приводящие борозды могут быть расположены на поверхности тела или на ловчих щупальцах. Первый случай представляют служащие для проведения пищи ко рту амбулакральные борозды некоторых морских звезд из отряда Phanеrozonia, Ко второму типу относятся, например, придатки губных щупалец Nuculidae и Nuculanidae (Protobranchia) (рис. 74, А);сюда же относится ловчий аппарат полихет сем. Тегеbellidae, образованный многочисленными длинными и тонкими щупальцами, подковообразно окружающими спереди рот.
4. Реснички расположены в приводящих бороздках, обращенных в толщу воды, улавливающих и приводящих ко рту пищевые частицы. Мерцательный аппарат такого рода чрезвычайно характерен для Pelmatozoa. Его возникновение и эволюция в основном определяли собой начальные этапы эволюции всех иглокожих. Морские лилии, нашедшие наиболее совершенный путь создания аппарата приводящих борозд, этим самым получили преимущество перед всеми остальными Pelmatozoa и одни дожили до наших дней. Заметим, впрочем, что в ловле пищи у морских лилий принимают участие и амбулакральные ножки. 5. Засасывающий мерцательно-вододвижущий аппарат свойствен губкам, плеченогим, двустворчатым моллюскам, некоторым Prosobranchia и ряду низших ордовых. Во всех этих случаях мерцательные эпителии располагаются внутри каналов, являющихся струенаправляющими приспособлениями, и создают в них токи воды, приносящие пищевые частицы к тому или иному улавливающему аппарату; наличие каналов, придающих определенное направление приносящим и уносящим токам, чрезвычайно увеличивает эффективность работы мерцательных эпителиев, в силу чего рассматриваемый тип вододвижущего аппарата является наиболее совершенным. Наиболее примитивные двустворчатые по способу питания являются собирателями донных отложений, используя в качестве орудия собирания длинные придатки околоротовых лопастей, снабженные мерцательными желобками (например, у Yoldia и других Protobranchia). Жабры их сохраняют одну дыхательную функцию и имеют строение типичных ктенидиев с короткими треугольными лепестками, и мерцательный покров жабр служит только для обновления воды около их поверхности, создавая для этого в мантийной полости лишь слабые токи воды. Ведущим фактором в эволюции двустворчатых явился переход от питания донными отложениями к питанию наннопланктоном, добываемым путем фильтрации; при этом ток воды, создаваемый ресничками жаберного эпителия, используется для привлечения пищевых частиц. В связи с этим происходит гипертрофия ктенидиев, лепестки которых разрастаются в длинные нити и нередко частично или полностью утрачивают дыхательную функцию. Вододвижущий аппарат пластинчатожаберных, в его наиболее совершенном виде, у Eulamellibranchia, устроен следующим образом. Основной движущей силой является ресничный эпителий прежде всего жабр и затем околоротовых лопастей. И те, и другие расположены в мантийной полости; края мантии в типичном случае либо плотно прикладываются друг к другу, либо срастаются. В итоге мантийная полость фактически открывается наружу всего лишь двумя рядом расположенными сифональными отверстиями.
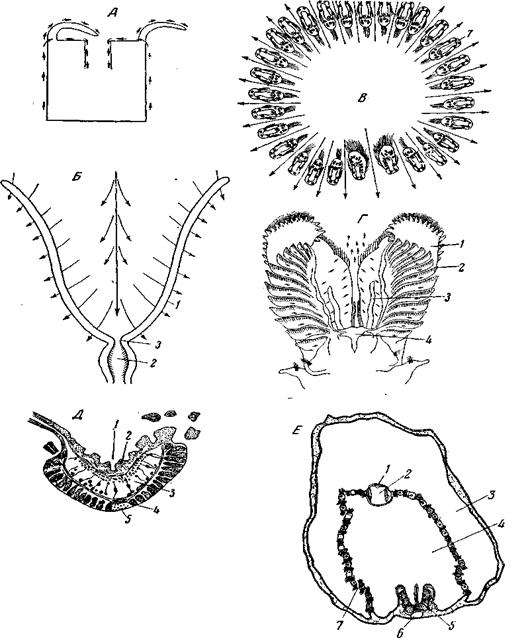
Рис. 73. Некоторые типы мерцательно-гидрокинетического аппарата (кроме моллюсков) А – актиния Protanthea simplex, схема токов воды на поверхности тела, вызываемых работой ресниц эктодермы (из Pax); Б – Bryozoa Gymnolaemata, схематический продольный разрез через венец щупалец, показывающий направление токов воды; 1 – щупальца; 2 – глотка; 3 – рот (по Borg); В – Flustrella (Bryozoa, Gymnolaemata), поперечный разрез через венец щупалец вблизи основания (по D. Atkins, из Hyman); Г – Sabellaria spinulosa (Polychaeta, Spiomorpha, Hermellidae), передний конец тела с вентральной стороны. Стрелками показаны направления токов воды, обусловленных работой ресниц; 1 – торчащие вперед лопасти, образованные сегментами проторакса и служащие ловчим аппаратом; 2 – ряды щупалец, сидящих на этих лопастях и покрытых мерцательным эпителием; 3 – пальпы; 4 – рот (по К.Е. Johansson); Д – Cephalodiscus nigrescens (Graptolithoidea), поперечный разрез через руку: Третье отверстие, через которое высовывается нога, часто обращено в грунт и так или иначе с гидравлической точки зрения не дает себя знать. Наружный жаберный листок прирастает краем к мантии, внутренний – к ноге; позади ноги оба внутренних жаберных листка срастаются между собой. Благодаря этому вся мантийная полость, за исключением ее передней, околоротовой части, оказывается разделенной на два отдела – спинной и брюшной. Работа ресниц гонит воду по брюшной части мантийной полости вперед, ко рту; оттуда по спинной части вода идет назад, мимо мочевого, полового и анального отверстий, к выводящему сифону. Таким образом, мантийная полость представляет собой U-образно изогнутый канал, оба отверстия которого, засасывающее и выводное, расположены на заднем конце животного, у роющих форм торчащем из грунта. По всему пути вдоль жабр вода понемногу профильтровывается между жаберными нитями и проходит в спинной, выводной отдел мантийной полости. При этом пищевые частицы задерживаются на жабрах и вдоль этих последних постепенно перегоняются кпереди, к губным щупальцам и оттуда – ко рту, частично – прямо ко рту. Благодаря громадной поверхности мерцательного эпителия жабр и упорядочению создаваемых им токов, работа ресниц обеспечивает значительную сосущую силу, и пластинчатожаберные принадлежат к числу наиболее совершенных фильтраторов. Как строение жабр, так и детали устройства и расположения ресничек достигают у высших пластинчатожаберных высокой степени совершенства. Ротовые лопасти, или пальпы, несут у большинства пластинчатожаберных функцию сортировки поступающих на их поверхность частиц: мелкие частицы направляются в рот, более крупные – сбрасываются в мантийную полость и в дальнейшем выводятся прочь через выводной сифон, благодаря перистальтике последнего (см. рис. 74, В),или, при его отсутствии, – благодаря резкому хлопанию створок раковины, как у Pectinidae. Надсемейство Tellinacea (Eulamellibranchia) вторично вернулось к питанию донными отложениями, имея уже совершенный гидрокинетический аппарат и хорошо развитые сифоны (рис. 74, В). В этом случае для собирания донных отложений животное приближает к их поверхности свой вводной сифон, и верхний слой ила (пелоген) втягивается в мантийную полость силой тока воды, создаваемого жаберными ресничками. Другими словами, в соответствии с законом необратимости эволюции, при возвращении высших Lamellibranchia к питанию донными отложениями, прежде служившие для этого органы не восстанавливаются, и прежняя функция осуществляется новым путем. Среди улиток использование мерцательно-вододвижущего аппарата для привлечения пищи независимо возникло по меньшей мере в пяти различных семействах переднежаберных: у сидячих Calyptraeidae Vermetidae и Capulidae и у свободноподвижных Turritellidae и Struthiolariidae
Рис. 74. Некоторые типы мерцательно-гидрокинетического аппарата моллюсков А – Yoldia limatula (Bivalvia Protobranchia), собирающая наилок при помощи придатков ротовых лопастей: 1 – вводной сифон; 2 – выводной сифон; 3 – сифональные щупальца; 4 – снабженные собирательной бороздой щупальца ротовых лопастей (по Lang); Б – Saxicava arctica (Eulamellibranchia), вид с брюшной стороны: 1 – раковина; 2 – выводной сифон; 3 – вводной сифон; 4— сросшаяся мантия; 5 – переднее отверстие мантии; 6 – нога (из Pelseneer); В – Tellina tenuis (Eulamellibranchia), вид справа, после удаления правой створки раковины и правой лопасти мантии: 1 – ротовые лопасти; 2 – передний мускул-замыкатель; 3 – рот; 4 – край мантии; 5 – нога; 6 – внутренний листок правой жабры; 7 – желоб мантийной полости, отводящий кзади отброшенные ротовыми лопастями частицы; 8 – комок таких частиц (в дальнейшем они выводятся наружу перистальтическими движениями вводного сифона); 9 – перекрещенные мышцы мантии; 10 – вводной сифон; 11 – выводной сифон; 12 – задняя кишка; 13 – задний мускул-замыкатель; 14 – задний ретрактор ноги; 15 – наружный листок жабры, стрелки указывают направление токов воды (по Yonge); Г – Struthiolaria papulosa (Prosobranchia Taenioglossa Strombacea), в нормальном положении зарывшаяся в грунт, успевшая проделать ход для выходящего тока и проделывающая хоботом отверстие для входящего тока; 1 – сильно вытянутый хобот; 2 – нога; 3 – крышечка; 4 – канал для выходящего тока, проделанный в грунте; Д – Struthiolaria, вынутая из раковины, мантийная полость вскрыта вдоль ее правой стороны; Е – она же, схематичный поперечный разрез через мантийную полость; Во всех этих случаях происходит смена функции, и для привлечения пищи начинает использоваться мерцательный покров ктенидиев, первоначально служащий лишь для обновления воды вокруг жабр. В крайних случаях (у Calyptraeidae) ктенидии в значительной мере утрачивают дыхательную функцию, переходящую к стенкам мантийной полости, как это имеет место и у многих Lamellibranchia. При этом, как и у двустворчатых, лепестки ктенидиев вытягиваются в длинные нити, что служит к усилению их работы по созданию тока воды.
Поиск по сайту: |