|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Меиамерия других хелицеровых
Наземные хелицеровые, или паукообразные, отличаются значительной древностью: многие отряды паукообразных известны уже с карбона (Теlyphones, Araneina – подотряд Mesothelae, Solifugae, Ricinulei, Opiliones, а также вымершие Anthracomarti, Kustarachnida и др.); клещи отряда Acariformes известны даже с девона. Общее число сегментов тела в различных отрядах паукообразных различно. Среди Araneina Heptathela kimurai имеет 12 тергитов опистосомы (не включая прегенитальный сегмент), что соответствует полному, как у скорпионов, числу сегментов тела. Ни у кого другого из паукообразных оно не достигает полного числа – 19 сегментов, свойственного скорпионам (придаток анальной лопасти в счет, конечно, не входит). У Telyphones, Amhlypygi, Schizopeltidia, Pseudoscorpionoidea и Araneina имеется по 18 сегментов, у Palpigradi, Solifugae и Opiliones – 16 сегментов, у Ricinulei, Opilioacarina и Parasitiformes – 15 и, наконец, у Acariformes – 13 сегментов. Таким образом, у Acariformes вся метасома полностью редуцирована. Мало того, у некоторых представителей последнего отряда в результате педоморфоза (сохранения личиночных признаков у взрослой формы) число это уменьшается еще значительнее, и, например, Tydeidae (подотряд Prostigmata) имеют всего 10 сегментов. Но и среди Araneina 18 сегментов у взрослого животного хорошо выражены только в подотряде Mesothelae (сем. Liphistiidae) (рис. 40, Б, В).У высших Araneina 6 задних сегментов испытывают частичную редукцию – их вентральные части не развиты. В целом, мы видим у паукообразных тенденцию к укорочению тела путем редукции задних сегментов, доходящую до полного исчезновения метасомы, а у части акариформных клещей – также 1–3 последних сегментов мезосомы. У многих групп последние сегменты метасомы, еще не исчезнув, приобретают рудиментарный характер, представляя собой маленький постабдомен из сильно суженных и укороченных сегментов. Такой постабдомен свойствен Telyphones, Schizopeltidia (рис. 40, А;рис. 44, Б), а. также Palpigradi и Ricinulei (рис. 46, 3); во всех этих группах он состоит из двух сегментов и анальной лопасти. Частичный анаморфоз, который мы видели у эвриптерид, из наземных групп встречается только у Acariformes; например, в очень яркой форме – у Packygnathus. Личинка P. dugesi имеет 3 мезосоматических сегмента и анальную лопасть, протонимфа – 4, дейтонимфа – 5 и, наконец, тритонимфа – 6 мезосоматических сегментов. Таким образом, с каждой линькой у P. dugesi прибавляется по одному сегменту. У Tydeidae анаморфоз выпадает, и они на всю жизнь сохраняют личиночное число сегментов вместе с рядом других личиночных признаков (педоморфоз). Своеобразную черту в метамерию всех хелицеровых вносит судьба последнего – 7-го сегмента просомы. Только у Limulus он несет рудиментарные конечности. В других отрядах он выражен в весьма различной степени, хотя всегда слабее окружающих сегментов и никогда не несет конечностей (см., однако, выше: о метастоме эвриптерид).
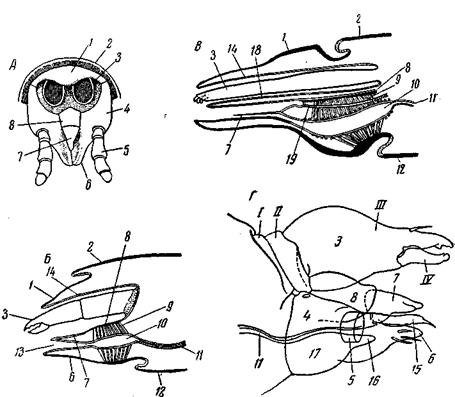
Рис. 40. Расчленение тела наземных Chelicerata А – Trithyraeus Cambridge! (Schizopeltidia), самка со спинной стороны: 1 – хвостовая нить (flagelum); 2 – метапельтидий; 3 – ямки на тергитах опистосомы – места прикрепления дорсо-вентральных мышц; пдп – коксы педипальп; хл – хелицеры (по Вorner); Б – Liphistius malayanus (Araneina, Mesothelae), видны тергиты опистосомы (по Abraham); Лучше всего он развит у Palpigradi (рис. 44, А),где представляет настоящий, обособленный, хотя и узкий, сегмент, служащий соединительным стебельком между просомой и мезосомой. У телифонов, схизопельтидий и пауков он тоже образует стебелек и снабжен тергитом истернитом Основной принцип специализации конечностей по тагмам, установившийся у водных хелицеровых, остается в силе и у наземных отрядов, но с некоторыми отклонениями. Конечности просомы остаются ротовыми и локомоторными, конечности мезосомы – дыхательными. Однако участие конечностей в принятии пищи сильно ограничено. Тогда как у мечехвостов рот окружен коксальными отростками всех просоматических конечностей, кроме хелицер, у современных скорпионов в образовании предротовой полости участвуют педипальпы и 2 пары ног, у Opiliones – педипальпы и
Рис. 41. Ротовые органы хелицеровых (кроме клещей) и сравнение передней части тела хелицеровых с головой ракообразного (по Snodgrass) А – схема сегментации просомы паукообразного, пунктированы головная лопасть и ее производные; Б – паукообразное, вид спереди (схема), нижняя губа представляет собой стернит сегмента педипальп, как у Araneina; В – то же, после удаления хелицер; ротовые части срезаны позади пальп; Г – Xiphosura polyphemus, передний край рта; Д – схематический продольный разрез через Б; Е – Talorchestia longicornis (Crustacea, Amphipoda), голова спереди; антенны занимают сходное положение с хелицерами паукообразного 1 – основания педипальп; 2 – предротовая полость; 3 – верхняя губа (labrum);
Коксальные эндиты педипальп у этих последних покрыты волосками, образующими фильтр, который задерживает твердые частицы пищи и не допускает их в ротовую полость; после окончания еды скопившиеся на фильтре частицы счищаются работой других конечностей. Такую же роль предротового фильтра несут коксэндиты и у некоторых других паукообразных. В деталях строения ротовой аппарат паукообразных представляет замечательное разнообразие: почти в каждом отряде он построен по Для части клещей (Parasitiformes, Opilioacarina) весьма характерно появление обособленного отдела тела, несущего ротовые органы и обозначаемого как головка (capitulum), или гнатосома. Головка включает переднюю часть головной лопасти, иногда с частями двух первых сегментов тела – сегментов хелицер и педипальп; топографически заднедорсальная часть головной лопасти, несущая глаза, остается вне головки. Обособленность головки вытекает из двух особенностей (R. Snodgrass, 1948), из коих первая – склеротизация ее дорсальной стенки над основанием хелицер, т.е. образование tectum capituli (рис. 42, А–Б).На боках tectum срастается с заднедорсальными краями кокс педипальп, которые, в свою очередь, срастаются между собой на вентральной стороне тела; в силу этого основание головки образует сплошное склеритное кольцо – basis capituli. Вторая ее особенность в том, что головка, а именно basis capituli, тонкой перепонкой подвижно сочленена с остальным туловищем и обычно может втягиваться в углубление, называемое камеростомом, и высовываться из него действием протракторов. Телоподиты педипальп образуют свободные щупальца. Под основанием хелицер коксы педипальп соединяются между собой при помощи эпистома, который, как и у других паукообразных, несет на своем переднем крае верхнюю губу. На вентральной стороне коксы педипальп образуют вырост (гипостом), представляющий дно предротовой полости, крышу которой образует верхняя губа. Что касается Acariformes, у них настоящего capitulum, по-видимому, не бывает; их хелицеры либо вообще остаются свободными, либо срастаются с коксами педипальп (у Cheyletidae), но во всяком случае не лежат в трубке, образованной срастанием кокс пединальп с tectum capituli. Однако при всем многообразии строения ротовых органов клещей весьма характерным является непременное участие в их образовании коксоподитов педипальп. В этом отношении клещи принципиально сходны с большинством паукообразных. Итак, возрастающая гетерономность просоматических конечностей паукообразных сводится к постепенному уменьшению числа конечностей, участвующих своими коксоподитами в принятии пищи, причем дольше всего эту функцию сохраняют педипальпы. Полностью она выпадает только в отряде Palpigradi. Заметим, что по номеру сегмента, к которому они принадлежат, педипальпы являются гомологами мандибул ракообразных, насекомых и многоножек.
Рис. 42. Ротовые органы клещей А – схема головки (capitulum) Parasitilorrnes, хелицеры удалены; Б – схематический продольный разрез через переднюю часть тела паразитиформного клеща; В – то же, в частности – Ixodoidea (по Snodgrass); Г – ротовой аппарат низших Acariformes (полусхематично, вид с правой стороны, телоподиты педипальпов отрезаны). Обратить внимание на свободные, 4-члениковые хелицеры (по А.А. Захваткину) 1 – tectum capituli; 2 – спинной щит (сагарах); 3 – хелицеры (на рис. А – места их прикрепления); 4 – коксы; 5 – телоподиты педипальп; 6 – коксэндиты; 7 – верхняя губа (labrum); 8 – эпистом; 9 –дорсальный дилататор глотки; 10 – глотка; 11 – пищевод; 12 – стернит III сегмента просомы (тритостернум); 13 – рот; 14 – верхняя стенка влагалища хелицер; 15 – придатки коксальных зндитов педипальп, так называемые максиллы;
Что касается конечностей мезосомы, то для всего класса паукообразных характерна тенденция к их последовательной редукции. Столь полного набора мезосоматических конечностей, какой свойствен скорпионам, мы больше не находим ни в одном из отрядов. Половых крышек на 8-м сегменте больше нет ни у кого, если не считать части Opiliones. У Araneina на заднем крае этого сегмента располагаются легочные отверстия; можно было бы думать, что эта передняя пара легочных мешков принадлежит следующему, 9-у сегменту, а сидящая позади него задняя пара легочных мешков – 10-у. Однако 10-й сегмент имеет собственные конечности в виде первой пары паутинных бородавок. Таким образом, приходится допускать, что вторая пара легочных мешков Araneina принадлежит 9-у сегменту, а первая пара – 8-у, половому, сегменту и гомологична, таким образом, половым крышечкам (operculum) мечехвоста. Легкие телифонов и фринов расположены на 8 и 9-м сегментах, единственная пара легких схизопельтидий лежит на 8-м сегменте. Одинаковое расположение легких у телифонов, фринов и пауков подтверждает близость этих отрядов между собою. 9-й сегмент, у скорпионов несущий гребневидные органы, у большинства отрядов лишен конечностей и только у телифонов, фринов (Amblypygi) и части Araneina несет вторую пару легочных мешков, а у других Araneina – пару трахей, гомологичную, как принято думать, легочным мешкам; у Solifugae он тоже несет пару трахей, но гомологичны ли эти последние легочным мешкам, сказать трудно. 10-й сегмент несет трахеи у лжескорпионов и бихорок и 1-ю пару паутинных бородавок у пауков. Таким образом, из 6 пар мезосоматических конечностей у пауков остается не более 4 пар, у телифонов и фринов – 2 пары, у схизопельтидий – 1 пара и, быть может, у лжескорпионов – 2 пары (если их трахеи считать гомологами легких, что не доказано). Мезосоматические трахеи имеются, сверх того, у сенокосцев (1 пара) и бихорок (3 пары), но их гомология легочным мешкам менее вероятна. Palpigradi, Ricinulei и клещи подобного рода остатков мезосоматических конечностей, хотя бы предполагаемых, вовсе не имеют. Таким образом, наряду с тенденцией к уменьшению числа сегментов метасомы у паукообразных наблюдается вторая тенденция – к редукции мезосоматических конечностей. Теряя свои конечности, мезосома теряет свое отличие от метасомы, и определение границ между обоими этими отделами нередко становится возможным лишь на основании точного счета сегментов. Поэтому в теле наземных хелицеровых очень часто различают не три отдела, а всего два: просому и опистосому, причем опистосома представляет сумму всех наличных сегментов мезосомы и метасомы. Видоизменения метамерии наружного скелета паукообразных весьма многообразны, но некоторые общие тенденции улавливаются и тут. Наиболее примитивный скелет членистоногого, как мы выше видели, носил бы гомономный характер и был бы представлен метамерно и гомономно расположенными наборами многочисленных склеритов. Наиболее примитивный скелет, фактически наблюдаемый, обычно уже несколько уклоняется от этого типа благодаря своей меньшей гомономности и взаимному слиянию одних склеритов и редукции или разрастанию других. Рассмотрим развитие гетерономности скелета членистоногих на примере Chelicerata.
У всех водных хелицеровых все тергиты просомы сливаются в один общий щит – карапакс (сагарах). В этом отношении многие из наземных паукообразных сохранили более примитивное строение, больше приближающееся к расчленению скелета трилобитов. Действительно, у ряда паукообразных, вместо сплошного щита, одевающего всю просому, имеется щиток – пропельтидий (propeltidium), покрывающий только ларвальный отдел тела, т.е. сегменты хелицер, педипальп и двух первых пар ног. Как полагает P. Schulze (1937), покрытый таким щитком отдел туловища вполне гомологичен голове трилобита, и пропельтидий более или менее гомологичен головному щиту трилобита. У Palpigradi и Schizopeltidia вслед за пропельтидием идет разбившийся на две парные пластинки мезопельтидий (тергит сегмента ног III), затем – метапельтидий (тергит сегмента ног IV) и затем ряд свободных и не слитых между собой тергитов опистосомы (рис.40, А). У бихорок (Solifugae) за пропельтидием следуют две пары склеритов (arci anteriores и posteriores), быть может соответствующие в своей совокупности мезопельтидию; затем идет метамерный ряд тергитов, включающий во всяком случае метапельтидий и тергиты опистосомы (рис.40, Д).Таким образом, по степени самостоятельности своих тергитов эти три отряда паукообразных значительно примитивнее водных хелицеровых и стоят на уровне трилобитов. Наряду с этим большинство паукообразных – Scorpiones, Telyphones, Amblypygi, все Araneina, Pseudoscorpionoidea, Ricinulei – имеет сплошной щит, прикрывающий всю просому, как у мечехвостов и эвриптерид. Среди сенокосцев (Opiliones) в сем. Phalangidae (рис. 40, Е)также имеется общий просоматический щит, но у Gagrellinae на нем поперечными швами намечены границы двух последних тергитов, т.е. видны еще следы первоначального расчленения щита на про-, мезо- и метапельтидий. У других Opiliories с просоматическим щитом сливается еще несколько тергитов опистосомы, но границы этих последних так же, как и границы мезо- и метапельтидия, большей частью продолжают быть заметны в виде швов. Что касается тергитов мезосомы, они остаются самостоятельными у Scorpiones, Telyphones, Amblypygi, Schizopeltidia, Palpigradi, Solifugae, Pseudoscorpionoidea, Ricinulei, в примитивных группах Acariformes и среди Araneina – у примитивного сем. Liphistiidae (подотряд Mesothelae). У остальных Araneina склериты как таковые в пределах опистосомы вообще исчезают, вся опистосома покрыта мягкой кожей, и внешние проявления метамерии исчезают. Чрезвычайное разнообразие представляет расчленение спинной стороны тела у клещей.
Рис. 43. Расчленение тела у различных клещей А – Arthrosebaia palmacea (Acariformes Endeostigmata), со спины, трилобитоидное расчленение: I – пропельтидий; II – мезопельтидий; III – метапельтидий; 1–5 – видимые со спинной стороны сегменты мезосомы; Б – Speleorchestes poduroides (Endeostigmata), со спины; тритиреоидное расчленение; 1 – гнатосома (т.е. ротовой аппарат, образованный хелицерами и педипальпами); 2 – проподосома; 3 – метаподосома; 4 – опистосома (из А.А. Захваткина); В – Tyrolichua casei (Acariformes Tyroglypboidea), самец со спины; акароидное расчленение; ряды щетинок определяют последовательные сегменты гистеросомы (по А.А. Захваткину); Г – Microtrombidium hystricinum (Acariformes Parasitengona), никаких следов ни тагм, ни метамерии опистосомы (по Vitzthum); Д – Phyllocoptei triceras (Acariformes Tetrapodyli), самка сбоку; акароидное расчленение две задние нары ног редуцированы; вторичная метамерия постларвального тела, негомологичная метамерии зародыша (по Borner); Е – Rhodacarus (Parasitiformes Rhodacaridae), арахноидное расчленение; шунктиром показаны предполагаемые границы сегментов, установленные на основании изучения хетотаксии: I–VI – сегменты иросомы; VII – прегениталышй сегмент; VII – XIII – сегменты мезосомы; 1 – хелицеры; 2 – педипальпы; 3 – tectum capituli, заканчивающийся спереди копьевидным выступом; 4 – передний отдел спинного щита; 5 – задний отдел спинного щита; 6 – дорсальный щит мезосомы (оригинал А.Б. Ланге)
У Acariformes прежде всего мы встречаем расчленение, приближающееся к таковому бихорок, схизопельтидий, а в конечном итоге также и примитивных трилобитов, которое мы можем обозначить как трилобитоидное: головная лопасть и 4 первых сегмента, обозначаемые в совокупности как п р о т е р о с о м а, покрыты общим щитом (пропельтидием), тогда как 2 последних сегмента просомы и все сегменты мезосомы остаются свободными. Такой тип расчленения представляют, например, Pachygnathi-dae (Acariformes, Endeostigmata) (рис. 43, А).Далее мы встречаем тип расчленения, обозначаемый Захваткиным как тритиреоидный; тело разбито на три отдела: протеросому, состоящую из головной лопасти и 4 сегментов, Между первыми двумя типами существуют и переходные формы; например, первый наличный тергит мезосомы может остаться свободным, остальные сливаются между собой (Probrachychilwnius из Acariformes Oribatei). Из тритиреоидного типа путем слияния метаподосомы с одним из прилегающих крупных отделов выводятся два следующих типа расчленения: акароидный и арахноидный. Первый возникает в том случае, когда метаподосома сливается с метасомой в единую гисторосому (рис. 43, В). Этот тип расчленения свойствен очень многим Acariformes. Арахноидный тип расчленения возникает при слиянии метаподосомы с протеросомой, т.е. при полной интеграции просомы; среди клещей ему следуют Parasiliformes (рис. 43, Е).Спинной щиток (scutum) иксодовых клещей гомологичен, по P. Sclmlze (1937), просоматическому щиту (сагарах) пауков. Наконец у очень многих клещей происходит полное слияние всех отделов тела наряду с исчезновением метамерии скелета, и тело приобретает полностью интегрированную, иногда почти шарообразную форму (например, у многих Hydrachnida и других Acariformes Prostigmata; Замечательную особенность некоторых сенокосцев, например подотряда Cyphophthalmi, составляет загибание заднего конца тела на брюшную сторону. Внешне оно выражается в том, что 9 и 10-й опистосомальные тергиты смещаются у них на брюшную сторону. При этом 10-й тергит оказывается впереди 9-го; соответственно сдвинуто вперед и анальное отверстие, а стерниты 6, 7, 8 и 9-й изогнулись дугообразно, чтобы дать место передвинувшимся сюда тергитам (рис. 44, Д).Аналогичное явление в гораздо большем масштабе мы встречаем и у многих Acariformes (Oribatei, Tyroglyphoidea), у которых на брюшную сторону сместились тергиты трех последних сегментов мезосомы. Интеграция и укорочение тела путем загибания заднего конца на брюшную сторону свойственны и другим членистоногим, например, Crustacea Decapoda; однако у клещей и сенокосцев это загибание сопровождается редукцией брюшной стороны загнутых сегментов, т.е. ведет к значительно более высокой степени интеграции туловища, чем у десятиногих раков. Обращаясь к скелету вентральной стороны, мы прежде всего должны заметить, что у трилобитов стернитов нет. Между коксами ног трилобитов тянется узкая борозда, морфологически занимающая место шва замкнувшегося бластопора; по аналогии с листоногими раками можно полагать, что она служила для проведения ко рту пищи, подхватываемой эндитами туловищных ног. У хелицеровых, как мы видели, происходит последовательная редукция этих эндитов, сначала в области опистосомы, а затем и в области просомы. Вместе с этим исчезает и пищевая борозда, и может начаться склеротизация вентральной стороны тела. Эта последняя происходит за счет двух процессов: 1) образование метамерных или неметамерных (объединенных) стернальных щитов и 2) разрастание кокс и неподвижное прирастание их к брюшной стенке тела. Первый тип образования вентрального скелета в чистом виде наблюдается у Palpigradi. Причисляемый иногда сюда верхнеюрский Sternarthron zitteli имел отдельные стерниты во всех сегментах просомы и мезосомы. У современных Koeneniidae сегменты хелицер, трех задних пар ноги большинство сегментов опистосомы обладают самостоятельными стернитами, и лишь стерниты педипальп и ног I слились в одну общую пластинку (рис. 44, А), а стерниты сегментов постабдомена (метасомы) слились каждый со своим тергитом в цилиндрик. Второй тип образования вентрального скелета в чистом виде представлен в просоме сенокосцев, лжескорпионов, Ricinulei и ископаемых Kustarachnida; здесь брюшная сторона сплошь занята коксами ног, а стернитов нет вовсе или они представлены ничтожными рудиментами. В других отрядах мы встречаем различные промежуточые состояния. У телифонов (рис. 44, Б),наряду с увеличенными коксами, в просоме имеются три стернита. У Amblypygi и родственных им Araneina (рис. 44, В)ноги расставлены в стороны и между ними остается свободное пространство, занятое единым грудным щитом (sternum). Последний представляет результат склеротизации брюшных покровов, охватывающей область всех сегментов ходильных ног, кроме, может быть, последнего из них. Стернит сегмента педипальп у пауков хорошо развит и играет у них роль нижней губы. В области опистосомы стерниты почти во всех отрядах развиты хорошо и независимы друг от друга; отсутствуют они только у Araneina и во всех отрядах клещей; у ряда клещей вентральная поверхность опистосомы несет парные и непарные щитки неметамерной природы. У скорпионов, сенокосцев и Rici-nulei наблюдается сильный сдвиг передних стернитов опистосомы кпереди, особенно резко выраженный у сенокосцев (рис. 44, Д). Благодаря этому сильно сдвигается кпереди и половое отверстие. Во всех трех отрядах этот сдвиг совершается за счет гипертрофии ближайших постгенитальных стернитов (9 и отчасти 10-го) и влечет за собой полную редукцию стернита прегенитального (7-го) сегмента и значительное уменьшение стернита (8-го) полового сегмента. У сенокосцев происходит в то же время слияние стернитов 9 и 10-го сегментов. Итак, рассмотрение метамерии скелета паукообразных показывает, что весьма мало вероятно происхождение всех отрядов паукообразных от известных нам водных хелицеровых: первичных скорпионов, эвриптерид и мечехвостов. У многих из современных паукообразных мы находим метамерию, значительно более примитивную, приближающуюся к таковой трилобитов, а у Acariformes имеет место даже частичный анаморфоз. Более вероятным является независимое происхождение отдельных групп наземных хелицеровых от различных групп водных хелицеровых, из которых большая часть нам пока неизвестна. Однако можно считать очень вероятным, что все группы наземных хелицеровых исторически произошли все-таки от водных хелицеровых, а не непосредственно от трилобитов. В пользу этого положения, помимо редукции антенн I, говорит постоянство числа сегментов просомы и мезосомы и, в особенности, рудиментация 7-го сегмента тела у всех хелицеровых. Надо заметить, что в других группах членистоногих подобная редукция сегментов, лежащих на сгибе между двумя тагмами, хотя и встречается, но далеко не с тем постоянством, которое мы видели здесь. Таким образом, при допущении полифилетического происхождения хелицеровых пришлось бы считать наличие рудиментарного сегмента на сгибе двух тагм у всех представителей этой группы, в особенности же тот факт, что этот сегмент всегда является 7-м по общему счету, – трудно объяснимым совпадением. Подводя итоги, мы можем сказать, что эволюция метамерии в ряду от трилобитов к высшим хелицеровым сводилась к следующим изменениям: 1) дифференцировка трех отделов тела, или тагм, характеризуемых специализацией их конечностей или редукцией этих последних;
Рис. 44. Метамерия скелета вентральной стороны наземных Chelicerata А – Koenenia mlrabilis (Palpigradi), просома самки; Б – Telyphonus caudatus (Telyphones), самка; хвостовая нить изображена лишь частично (по Воrnеr); В – Heptatela kimurai (Araneina Mesothelae) (no Kashida); Г – Chelifer cancroides (Pseudoscorpionoidea), самец (по Borner); Д – Stylocellus (Opiliones Cyphophtlialmt) (no Hansen and Sorensen) 1 – рот; 2 – верхняя губа; 3 – паутинные бородавки; 4 –вертлуг первой пары ног; 5 – слившиеся стерниты третьего и четвертого сегмента просомы; 6 – постабдомен;
во-первых, к смещению головной лопасти вдоль спинной стороны кзади и всего сегмента хелицер на спинную сторону тела и, во-вторых, к смещению тергитов последних сегментов опистосомы на брюшную сторону со сдвигом анального отверстия по брюшной стороне кпереди и с редукцией соответствующих стернитов; 8) в ряде случаев – далеко идущая общая интеграция тела. В результате этой эволюции наиболее специализованные хелицеровые, вроде многих клещей, обладают коротким, компактным телом, интеграция всех метамер которого повела к почти полной редукции метамерии, а будто бы упрощенная архитектоника скрывает сложные перестройки, связанные с изгибанием оси метамерии.
Вопросы для самоконтроля
1. Характеристика метамерии трилобитов. 2. Строение конечностей трилобитов, хелицеровых и ракообразных. 3. Наружная морфология скорпионов. 4. Расчленение тела других хелицеровых. 5. Расчленение тела у различных клещей. 6. Метамерия скелета паукообразных. 7. Вопросы происхождения наземных паукообразных. 8. Эволюция метамерии хелицеровых. ТЕМА 7
МЕТАМЕРИЯ РАКООБРАЗНЫХ
7.1. Метамерное строение головы ракообразных
Среди ракообразных наиболее примитивный план строения и, в частности, наиболее гомономную метамерию сохранили Anostraca, во многих отношениях являющиеся примитивнейшими из всех членистоногих вообще. Однако наряду с этим примитивные черты, отсутствующие у Anostraca, разбросаны по другим группам ракообразных. Низшие раки – очень древняя группа. В кембрии из ее представителей уже существовал ряд форм, примыкающих к различным группам Phyllopoda, а также формы, быть может соединяющие признаки Phyllopoda Conchostraca с таковыми Ostracoda. При сравнении Anostraca и Phyllopoda с трилобитами мы видим, что каждая из этих групп сохранила некоторые черты гомономной метамерии, отсутствующие в другой группе. Трилобиты отличаются более гомономной метамерией своих конечностей, зато процесс цефализации зашел у них очень далеко, ибо, как мы видели, всем трилобитам свойственна голова, включающая по меньшей мере четыре постантеннулярных сегмента. Наоборот, у Phyllopoda Anostraca (рис. 45, А)в состав головы входит всего-навсего один постантеннулярный сегмент; зато четыре первые пары постантеннулярных конечностей резко специализованы и различаются как между собой, так и по сравнению с остальными туловищными ногами. Туловище ракообразных также никогда не достигает той степени гомономности, которую мы встречаем в примитивных семействах трилобитов. В подклассе Malacostraca обычно все сегменты тела несут конечности, как у трилобитов, однако в отличие от трилобитов, передний отдел туловища высших раков (грудь) значительно отличается от заднего отдела (брюшка) как по форме своих сегментов, так и по направлению специализации их конечностей. У Entomostraca[2] туловище обычно содержит по меньшей мере две тагмы: грудь и брюшко, из которых последнее, подобно метасоме хелице-ровых, лишено конечностей. Однако у Conchostraca все сегменты тела, вплоть до анальной лопасти, несут конечности, которые лишь постепенно уменьшаются в размерах к заднему концу (рис. 47, В),так что говорить здесь о наличии обособленного брюшка не приходится. Lipostraca
Рис. 45. Метамерное строение головы ракообразных А – Eubranchipus vernalis (Phyllopoda Anostraca), протоцефалон в виде обособленной головы, мандибулярный сегмент (II) независим, максиллярные (III и IV) – слиты между собой; Triops longicaudatus (Phyllopoda Notostraca): Б – со спинной стороны; челюстные сегменты слиты с протоцефалом, максиллярный тергит вытянут в большой щит; В – с брюшной стороны; Г – Daphnia pulex (Cladocera): состав головы, как у Triops, тело покрыто двустворчатой раковиной, образованной за счет максиллярных тергитов; Д – Anaspides tasmaniae (Syncarida) протоцефалон с придатками, вид спереди; Е – Spirontocaris polaris (Decapoda Macrura), протоцефалон отпрепарован от панциря, с которым бывает подвижно сочленен; Ж – Callinectes sapidus (Decapoda Bracniura); протоцефалон с придатками, вид со спины; З – Chloridella panamerisis (Stomatopoda) протоцефалон, отпрепарованный от панциря, вид со спинной стороны; И – Orcttestoidea californica (Ampuipoda), голова (синцефалон) сбоку 1 – протоцефалон; 2 – первые пары грудных ног; 3 – верхняя губа; 4 – сложные глаза; Все это показывает, что представление о наиболее примитивном типе метамерии членистоногих мы могли бы получить, лишь соединив примитивные черты, разбросанные в различных группах. Строение головы ракообразных проанализовано R. Snodgrass (1938), выводов которого мы и придерживаемся ниже при изложении этого вопроса. Голова бранхиподид (Anostraca, рис. 45, А)образована предротовой лопастью (включая в это понятие и область антенн I) и одним постантеннулярным сегментом, дающим начало тритоцеребральному ганглию. Конечности этого сегмента имеют у личинки (у науплиуса) обычную двуветвистую форму, свойственную конечностям ракообразных, и коксы их снабжены жевательными эндитами; у взрослого животного эти конечности специализуются в качестве второй пары антенн (антенны II). Голова Eubranchipus представляет широкую монолитную капсулу, несущую стебельчатые сложные глаза, две пары антенн и верхнюю губу. Последняя, подобно верхней губе трилобитов и хелицеровых, представляет собой опускающуюся над передним краем рта складку кожи. Голова Anostraca, состоящая из предротовой лопасти и одного сегмента, обозначается в анатомии ракообразных как первичная голова, или п р о тоцефалон (protocephalon). Вслед за головой лежит второй сегмент тела, представленный небольшим тергитом и несущий пару конечностей специализованных в качестве жвал, или мандибул. Тергиты третьего и четвертого сегментов тела сливаются в один; их конечности – максиллы I, или максиллулы, и максиллы II, или максиллы, – также специализованы у громадного большинства раков в качестве жевательных органов; однако у Hutchinsoniella macracantha (Cephalocarida) максиллы II ничем не отличаются от туловищных конечностей, а максиллы I отличаются от них меньше, чем у всех других раков. У Phyllopoda обе пары максилл рудиментарны. Мускулы обеих пар антенн, а также мускулы глаз и верхней губы прикрепляются у Phyllopoda к стенкам головной капсулы. Мускулы мандибул прикрепляются в пределах своего сегмента, так что места прикрепления мышц строго метамерны. Кроме Anostraca, самостоятельный протоцефалон свойствен всем высшим ракам (Malacostraca), за исключением Leptostraca, Amphipoda и Isopoda. Маленькая головка высших раков обычно бывает более или менее прикрыта сверху направленным кпереди выступом мандибулярного сегмента, так называемым рострумом (rostrum, рис. 45, Е).Она бывает подвижно сочленена с мандибулярным сегментом и несет глаза, обе пары антенн и верхнюю губу. У крабов (Decapoda Brachiura) эта голова очень мала (рис. 45, Ж)и запрятана в углубление под передним краем панциря (сагарах). У Stomatopoda (рис. 45, 3) она, наоборот, очень сильно развита и покровы ее распадаются на несколько метамерно расположенных склеритов; однако метамерия их не совпадает с эмбриональной метамерией, Из этого описания легко видеть, как неточны ходячие представления, согласно которым голова раков всегда состоит из 5 сегментов (включая сегмент антеннул), а у десятиногих сливается с сегментами груди в единую головогрудь. Утверждение о пятичленистой голове, свойственной всем ракам, основано лишь на том, что первые 5 пар конечностей более или менее сходно специализованы во всем классе ракообразных и что у многих ракообразных несущие их сегменты слиты в единую голову. Однако у десятиногих раков и ряда других форм челюстные сегменты в состав головы фактически не входят и эта последняя представлена одним протоцефалоном. Челюстные сегменты Decapoda входят в состав особой тагмы, свойственной этим последним, а именно челюстегруди (gnathothorax) (см. ниже). Всем прочим ракообразным свойственна гораздо более сложная голова, в состав которой, помимо первичной головы (protocephalon), входят по меньшей мере все сегменты челюстного отдела (gnathoce phalon). При этом максиллярная область головы очень часто образует щит или двустворчатую раковину в виде складки, покрывающей переднюю часть туловища (Notostraca, Leptostraca и другие), все туловище (Cladocera) или даже все тело (Conchostraca, Ostracoda). Образование это чрезвычайно характерно для ракообразных, являясь не только защитным приспособлением, но у многих форм также и струенаправляющим. У Notostraca тергиты первичной головы образуют переднюю часть спинного щита, часто отграниченную швом (рис. 45, В).Позади них хорошо заметен тергит мандибулярного сегмента, тогда как максиллярные тергиты, за счет которых образуется и свободная задняя часть щита, полностью слиты между собой, как у Eubranchipus. Щит Notostraca, подобно щиту трилобитов, образован за счет головной лопасти (включая область антенн I) и 4 первых сегментов. Однако в отличие от трилобитов, головная лопасть Notostraca лежит всецело на переднем краю щита и не образует кзади боковых продолжений, соответствующих свободным щекам трилобитов; вся задняя часть щита Notostraca всецело образована за счет максиллярных тергитов. Таким образом, несмотря на состав из тех же метамер, щит щитней представляет значительно более простое образование, с менее нарушенной метамерией, чем щит трилобитов или мечехвостов. Возможно, что состав щита трилобитов и щитней из одних и тех же метамер является простым совпадением; но весьма вероятно, что это сходство говорит о действительной близости обеих групп. Раковинные листоногие – Conchostraca и Cladocera (рис. 45, Г) – по строению головы в принципе сходны с Notostraca, однако головной щит загибается у них по бокам тела, что придает ему двустворчатую форму; кроме того, границы сегментов на щите исчезли и о задней границе У свободноживущих веслоногих (Copepoda) панциря в виде складки, прикрывающей туловище, не образуется, но все челюстные сегменты и первый постмаксиллярный сливаются между собой и с первичной головой (protocephalon) в одну крупную составную голову (syncephalon) (рис. 49, А).Конечности последнего сегмента этой головы принимают участие в работе ротового аппарата и обозначаются как челюстные ножки (максиллипеды). Голова веслоногих не представляет, однако, укороченного компактного целого. Это – просто слившиеся передние сегменты тела, сохраняющие форму простого участка метамерного туловища. Голова веслоногих не имеет подвижного сочленения с грудью. Голова синкарид (рис. 45, Д; 46) по уровню интеграции стоит не выше головы Copepoda. По составу она включает у Bathynellacea и ископаемых Gampsonychidea протоцефалон и гнатоцефалон, а у современных Anaspidacea, кроме того, и первый постмаксиллярный сегмент. Наиболее совершенной формой головы из всех ракообразных обладают Amphipoda и Isopoda. Голова бокоплавов и равноногих имеет такой же морфологический состав, как и голова Anaspidacea; она также включает протоцефалон, три челюстных сегмента и обычно один сегмент челюстных ножек; реже ногочелюстных сегментов бывает два или ни одного. Однако в отличие от головы веслоногих голова бокоплавов или равноногих представляет компактное, высокоинтегрированное целое, нередко подвижно сочлененное с грудью, напоминающее по внешнему виду голову насекомых (рис. 45, И). Это сходство побуждало некоторых авторов выводить насекомых из высших раков. Однако в действительности оно представляет лишь яркий пример конвергенции. У трилобитов и тех из ракообразных, у которых пища поступает в рот сзади (Phyllopoda, Cephalocarida, Copepoda, Euphausiacea, часть Mysidacea и другие – большей частью фильтраторы), в соответствии с этим рот нейтрален и направлен назад, labrum и антенны II тоже расположены вентрально, антенны I – вентрально или терминально; брюшная нервная цепочка у таких форм прямая, вплоть до deutocerebrum, и лишь protocerebrum несколько отклонен в дорсальную сторону. Другими словами, ось метамерии остается на своем переднем конце столь же прямой, как у аннелид. В отличие от этого, у тех из Malacostraca, которые перешли к хватанию пищи челюстями, в особенности у большинства изопод и части амфипод, рот направлен вперед и лежит почти терминально, функционально-передний конец тела образован наружным краем labrum, обе пары антенн значительно сдвинуты на спинную сторону, глаза дорсальны. В нервной цепочке самым передним из ганглиев оказывается tritocerebrum, тогда как deuto- и protocerebrum загнуты дорсально и каудально, так что лежат над- и несколько позади тритоцеребра. Таким образом, изменение в способе принятия пищи повело к изменению в направлении рта, к сдвигу вперед labrum и ротовых конечностей, к сдвигу на спинную сторону антенн, к искривлению мозга. Напомним про такой же процесс искривления оси метамерии в передней части тела наземных хелицеровых; он имеет место и у многих насекомых, независимо возникая во всех трех классах.
Поиск по сайту: |