|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Метамерия груди и брюшка низших ракообразных
Туловище ракообразных, как уже упомянуто, распадается на два отдела: грудь и брюшко. Исключение из этого правила составляют только Euphyllopoda Conchostraca, туловищные сегменты которых лишь постепенно уменьшаются к заднему концу тела и все несут одинаковые листовидные ножки. Однако не вполне исключена возможность, что отсутствие у Conchostraca абдомена является следствием его редукции. Листоногие обладают большим числом сегментов тела и притом различным в различных группах; в этом отношении они являются наиболее примитивными из раков. У Notostraca общее число сегментов (включая голову) обычно больше 40, у Conchostraca – от 14 до 36, у Anostraca – от 23 до 31, у девонских Lipostraca – 23. Другая весьма примитивная группа современных ракообразных – Cephalocarida (рис. 50) – имеет всего 13 пар постантеннулярных конечностей, включая головные, и 10 безногих сегментов; таким образом, по общему числу сегментов они сходны с Lipostraca и многими Anostraca, но число туловищных конечностей у них меньше, а число безногих сегментов больше, чем у этих последних. У большинства Anostraca грудь состоит из 13 сегментов, из которых 11 передних несут по одной паре одинаковых листовидных ножек, тогда как конечности 12 и 13-го сегментов превращены в половые придатки;
Рис. 46. Голова Anaspides tasmaniae (Syncarida, Anaspidacea) в профиль, включен также первый сегмент груди 1 – глаз; 2 – антенна I; 3 – антенна II; 4 –щупик (телоподит) мандибулы;
Девонский Lepidocaris rhiniensis (рис. 47, Д) – представитель вымершего отряда Lipostraca – сходен с Anostraca отсутствием щита или раковины, общим расчленением тела и отроением ротовых придатков; подобно большинству Anostraca, он имеет 11 пар локомоторных конечностей. За ними следуют гоноподии и дальше брюшко. Общее число сегментов туловища равняется 19, тельсон заканчивается вилкой. Двуветвистые, плавательные антенны II напоминают таковые Conchostraca и Cladocera. Главное отличие от остальных Phyllopoda заключается в том, что только 3 передние пары ног – листовидные, остальные 8 пар несколько напоминают двуветвистые плавательные ноги Copepoda (Scourfield, 1926, 1940). В целом, форма эта не более примитивна, чем современные листоногие, однако имеет и примитивные признаки, например присутствие рудиментарных конечностей на первом и последнем сегментах абдомена. У Notostraca и Conchostraca половое отверстие всегда помещается на 11-м сегменте туловища, конечности которого видоизменены, но меньше, нежели конечности половых сегментов Anostraca. За половым сегментом следует еще ряд сегментов, несущих листовидные ножки. У Conchostraca ножки сидят, как мы видели, на всех сегментах туловища. Общее число этих последних, в отличие от Anostraca, колеблется даже в пределах отдельных видов. Чрезвычайно своеобразна метамерия туловища Notostraca. Если первые 11 сегментов груди имеют и у них по одной паре ножек, то последующие 16–20 сегментов несут по несколько пар – от 2 до 6 на каждом сегменте, причем число пар ножек, приходящихся на один сегмент, увеличивается кзади, а размеры ножек уменьшаются. Подобная полиподия сегментов – явление совершенно исключительное. Ее обычно объясняют слиянием или неразделением сегментов. Помимо полиподии, постгенитальные сегменты груди щитней значительно отличаются от прегенитальных и по строению мускулатуры и иннервирующих ее периферических нервов. В силу всех этих различий карцинологи нередко считают грудью Notostraca и Conchostraca только прегенитальные сегменты, а все остальные относят к брюшку; однако, придерживаясь обычных критериев, приходится принимать, что в состав груди входят все снабженные ногами сегменты. При такой трактовке следует все же различать в составе груди два подотдела: переднегрудь, состоящую из 11 первых сегментов, и заднегрудь, включающую все остальные. Последние 4–15 сегментов тела щитней лишены ног и образуют брюшко, которое заканчивается анальной лопастью, с парными нитевидными церками. Отдельные виды Notostraca различаются между собой числом постгенитальных (многоногих) сегментов, числом ног и числом безногих абдоминальных сегментов; все эти величины ваоьируют в пределах каждого отдельного вида. Непостоянство числа сегментов тела в пределах вида – черта глубоко примитивная и среди членистоногих очень редкая; кроме Notostraca и Conchostraca, такое непостоянство свойственно лишь части трилобитов и многоножек. Итак, в теле Phyllopoda по строению конечностей мы можем различить 4 отдела: 1) антеннальный отдел (protocephalon), в состав которого входит акрон и один сегмент; конечности его имеют характер антенн; этот отдел самостоятелен у Anostraca и слит со следующим отделом у Conchostraca и Notostraca; 2) челюстной отдел (gnathocephalon) с чисто жевательными конечностями (частично рудиментарными); 3) грудь (thorax, или р а r e i о n) с конечностями неспециализованными, выполняющими все основные функции примитивной конечности членистоногого: локомоторную, дыхательную и хватательно-жевательную; часть грудных конечностей бывает изменена в половые придатки; грудь разделяется на два подотдела: прегенитальный (prothorax) и постгенитальный (m e t а t h о г а х); 4) брюшко (abdomen, или р 1 е о n), вовсе лишенное конечностей. Заканчивается тело анальной лопастью (telson). Очень важной примитивной чертой листоногих и цефалокарид является наличие жевательных придатков (эндитов) на всех парах локомоторных конечностей. Придатки эти участвуют в захвате пищи и перенесении ее ко рту. В этом отношении листоногие и цефалокариды сходны только с трилобитами. У Hutchinsoniella (Cephalocarida) эндиты на антеннах II сохраняются вплоть до сравнительно поздних стадий постэмбрионального развития (H. Sanders, 1957), тогда как у других ракообразных антенны II бывают снабжены эндитами только на стадии науплиуса. У всех других ракообразных, кроме Phyllopoda и Cephalocarida, жевательные эндиты сохраняются лишь на тех конечностях, которые специализовались в качестве челюстей (мандибулы, максиллы I и максиллы II) или челюстных ножек. Это обстоятельство создает весьма существенное различие между ракообразными и хелицеровыми. У большинства ракообразных захват и жевание пищи при помощи коксальных эндитов становится основной функцией определенных пар конечностей, утративших при этом локомоторную функцию; наоборот, у хелицеровых подобных конечностей, специализованных в качестве челюстей, не бывает: у водных хелицеровых в жевании участвуют гнатобазы всех локомоторных просоматических конечностей, у наземных эта функция гнатобаз утрачена или приобретает своеобразное выражение (например, у Parasitiformes). Тем не менее наиболее постоянной связью с ротовым отверстием у паукообразных обладают, как мы видели, педипальпы, по номеру своего сегмента являющиеся гомологами мандибул Crustacea. Как мы видели, число сегментов туловища у примитивных листоногих велико и непостоянно и эволюция в пределах класса ракообразных, так же, как и в ряду Trilobita – Chelicerata, идет по пути стабилизации и уменьшения их числа. Уменьшение числа сегментов тела является одним из путей интеграции метамерного тела и перехода от однообразно расчлененной, червеобразной цепочки сегментов к компактному, гетерономному, централизованному целому. Однако уменьшение числа сегментов является таким путем, который влечет в качестве побочных последствий упрощение строения, уменьшение морфогенетических возможностей и преждевременную специализацию. Такого рода преждевременная специализация исключает возможность далеко идущего прогрессивного развития, хотя вполне допускает большое многообразие в деталях.
Рис. 47. Расчленение тела Phyllopoda А – Artemiopsis stephensoni (Anostraca), самец, с вентральной стороны 1 – глаза; 2 – щит или раковина; 3 – антенны I; 4 – антенны II; 5 – верхняя губа; 6 и 7 – мандибулы; 8 – половые конечности; 9 – брюшко; 10 – вилка (первичная); 11 – вторичная вилка; 12 – тельсон; 13 – вентральная (пищепроводящая) борозда;
Уже среди Phyllopoda мы находим подобный ряд уменьшения числа сегментов и их стабилизации на низких цифрах, а именно – ряд Conch-ostraca – Cladocera. Среди Conchostraca Limnadiidae имеют 17–32 пары ног, большинство других семейств – 14–28 пар, но Lyncaeidae – всего 10–
Рис. 48. Cypridopsls vidua (Ostracoda), вид слева, часть левой створки удалена 1 – место отхождения створок от головы; 2 – науплиевый глаз; 3 – верхняя губа;
слиты между собой, и лишь у Leptodora оно состоит из 4 сегментов; если принять столько же сегментов брюшка и для других Cladocera, общее число постантеннулярных сегментов полифемид будет 12 – одна из крайних степеней олигомеризации среди членистоногих. Отряд Ostracoda (рис. 48) представлен конечными, высокоспециализованными формами другого аналогичного ряда уменьшения числа сегментов и укорочения тела. Тело их полностью закрыто двустворчатой раковиной. Раковина имеется уже у науплиуса – еще один, пример сдвига на более раннюю стадию первого появления органа, особенно сильно развитого у взрослой формы (правило Кейбеля – Менерта, установленное на примерах из эмбриологии позвоночных). Голова несет антеннулы и обычные 4 пары постантеннулярных конечностей. Большинство представителей имеет, кроме того, еще 2 пары грудных ноги, в некоторых семействах – так называемую чистилку, которая, быть может, также представляет видоизмененную пару конечностей. У Polycope обе пары торакальных ног редуцированы. Таким образом, максимальное число постантеннулярных конечностей равняется у Ostracoda 6 или 7 (считая чистилку), минимальное – 4 парам. Брюшко, так же, как и грудь, не расчленено; у самцов оно несет сложный пенис, который некоторые считают видоизмененной парой конечностей; P. Weygoldt (1960) считает, что пенис принадлежит сегменту чистилки. Определить число сегментов, входящих в состав тела Ostracoda, очень трудно, но во всяком случае оно сильно редуцировано. Остракоды – один из древнейших отрядов ракообразных, известных с кембрия. Большой отряд Copepoda (веслоногие) в отношении состава тела из сегментов также построен довольно однообразно, если оставить в стороне сильно видоизмененные паразитические формы. Свободноживущие веслоногие (рис. 49, А)имеют голову, состоящую из протоцефалона и 4 следующих сегментов (включая сегмент максиллипед), грудь, состоящую из Число и характер специализации конечностей являются для свободноживущих веслоногих более постоянными признаками, чем число свободных сегментов; у различных Copepoda мы наблюдаем разные степени слияния сегментов: сегментов груди с головой и друг с другом, абдоминальных сегментов – между собой. Наиболее измененные паразитизмом семейства веслоногих (Lernaeidae, Lernaeopodidae и другие) во взрослом состоянии почти полностью теряют метамерное строение. Веслоногие в ископаемом состоянии неизвестны. Представители небольшого отряда Branchiura (рис. 51) являются эктопаразитами рыб. Их голова и грудь покрыты общим щитом. Головные придатки личинки довольно типичны, но у взрослого все они сильно видоизменены в связи с паразитизмом. Грудь несет 4 пары двуветвистых плавательных ног. Нерасчлененное брюшко превращено в небольшой плавник, в выемке которого лежит furca. Соблазнительно считать плавательные ноги гомологами плавательных ног Copepoda, но тогда пришлось бы допустить полную редукцию не только 6, но и 1-й пары парейопод (максиллипеды веслоногих), в пользу чего никаких доказательств пока нет. Примитивный признак Branchiura – наличие щита (сагарах) и сложных глаз. Усоногие (Cirripedia) сильно видоизменены сидячим образом жизни. Часть усоногих подвергается еще большему видоизменению благодаря переходу к паразитизму. Известны с палеозоя. Свободноплавающие личинки усоногих проходят стадию науплиуса, несколько метанауплиальных стадий и стадию циприсовидной личинки (рис. 49, Б).
Рис. 49. Расчленение тела Copepodoidea А – схема свободноживущего веслоногого (Copepoda), самка, с брюшной стороны (из Kukenthal); Б – циприсовидная личинка Lepas fascicularis (Cirripedia) (из Мао Bride); В – Lepas, взрослая особь (по Сh. Darwin); Г – Aecothorax ophioctenis, самец (по В.Л. Вагину) ант I – антенны первые; ант II – антенны вторые; мд – мандибулы; мкс I – максиллы первые; мкс II – максиллы вторые; 1–6 – торакальные конечности; 7 – половое отверстие; 8 – вилка; 9 – брюшко; 10 – ротовой конус Ascothorax; 11 – кишечник; 12 – пенис; 13 – раковина; 14 – раковинная (максиллярная) железа; 15 – мускул-аддуктор раковины; 16 – цементная железа; 17 – парные глаза; 18 – науплиальный глаз
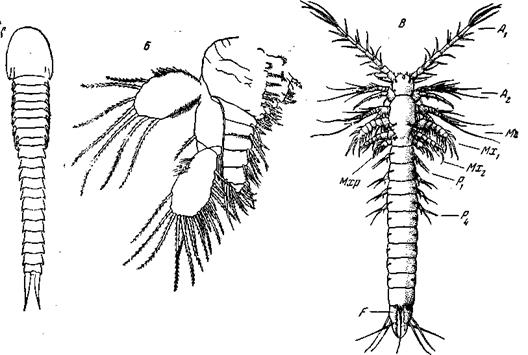
У Balanus crenatus бывает от 6 до 8 предциприсовых стадий. Циприсовидная личинка наличием и формой двустворчатой раковины напоминает взрослых Ostracoda, однако состав тела из сегментов и специализация конечностей отличны и напоминают веслоногих, а не остракод. Личинка усоногих имеет 2 пары антенн, мандибулы, 2 пары максилл и 6 пар грудных ножек. Всего постантеннулярных конечностей 10 пар. Брюшко рудиментарно, и число вошедших в состав его сегментов определить нельзя. Такая личинка, поплавав, прикрепляется головой к субстрату, антенны при этом редуцируются, и прикрепленная к голове раковинная складка образует мантию, в которой у типичных усоногих (рис. 49, В)и развиваются характерные для этих последних пластинки. Ротовые и грудные конечности у типичных форм (Pedunculata и Operculata) сохраняются полностью, у Acrothoracica число грудных уменьшено. Наконец у Rhizocephala (корнеголовых) никаких конечностей и вообще никаких следов метамерии у взрослого животного не остается. Итак, неопределенно большое число сегментов груди мы находим только среди Euphyllopoda. Во всех остальных рядах мы видим уменьшение и стабилизацию их числа, причем в различных рядах типичное число торакальных сегментов оказывается не совсем одинаковым: у Cladocera – 4–6, у Copepoda, Mystacocarida, Cirripedia и Ascothoracida – 6; то же число гипотетически можно приписать и Branchiura. Пять последних отрядов имеют много общего и могут быть объединены в один надотряд Copepodoidea; общим для них является сходное расчленение тела, а также торакальные конечности, построенные по двуветвистому, членистому типу,
Рис. 50. Наружная морфология Cephalocarida и Mystacocarida А, Б – Hutchinsoniella macracantha (Cephalocarida): A – вид с дорсальной стороны (конечности не врисованы); Б – одна из грудных конечностей
без жаберных и жевательных придатков. Исходное число торакальных конечностей для всех Copepodoidea равняется 6 парам; у Cirripedia и примитивных Ascothoracida они все специализированы одинаково, у Copepoda и Mystacocarida 1-я пара превращена в максиллипсды, а последняя у большинства копепод – в половые придатки, а у Mystacocarida – редуцирована. У Branchiura, если они принадлежат сюда, редуцированы и 1-я и последняя пары торакальных ног. Таким образом, в этих четырех отрядах состав груди, по-видимому, один и тот же, несколько различна лишь дифференцировка торакальных конечностей. Ostracoda и Phyllopoda сильнее отличаются как от них, так и между собой.
Поиск по сайту: |