|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Проявления диссимметрии у членистоногих
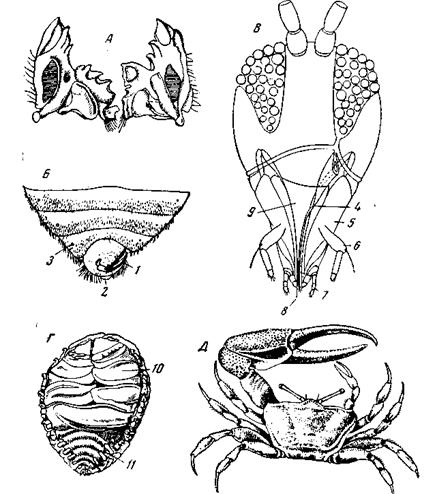
Среди Arthropoda проявления диссимметрии как во внутреннем строении, так и в наружном, многочисленны, в особенности в классах Crustacea и Insecta. Наоборот, среди трилобитов, пантопод, хелицеровых[3]и многоножек диссимметрия встречается редко. У насекомых часто встречаются мелкие нарушения симметрии, например, в строении мандибул (рис. 62, А), где они имеют тот же смысл, что и у Eunicidae – коаптация обеих мандибул к совместной работе. Существуют также случаи диссимметрии в строении мужских совокупительных органов у многих жуков, и т.д. По существу более значительную, хотя внешне мало заметную диссимметрию представляет задний конец брюшка самцов многих двукрылых, у которых два последних сегмента тотчас после выхода из куколки испытывают поворот вокруг продольной оси на 180°, так что их тергиты оказываются на брюшной стороне, а стерниты – на спинной. Диссимметричен конец брюшка у самцов водяного клопа Corixa, усамцов эмбий (Embioidea), у постельного клопа (Cimex) (рис. 62, 5); у последнего диссимметрия вызвана асимметрией симбиотического органа[4]. У трипсов (Thysanoptera) наблюдается резкая диссимметрия в строении головы, обусловленная редукцией одной из мандибул (рис. 62, В);на внешних очертаниях головы эта диссимметрия, однако, почти не отражается.
Рис. 62. Проявление диссимметрии у насекомых и ракообразных А – мандибулы Xylotrupes gideon, (Coleoptera) (по Berlese); Б – задний конец брюшка постельного клопа Сгтех lectularius (Hemiptera) с брюшной стороны; 1 – пенис; 2 – задний проход; 3 – дыхальца (по Martini); В – голова Aeolothrips fasciata (Tnysanoptera) со спинной стороны лба: 4 – единственная мандибула; 5 – максиллы I; 6 – щупальца максилл I; 7 – щупальца нижней губы (максилл II); 8 – парные колющие щетинки (внутренние лопасти первых максилл); 9 – верхняя губа (по Handlirsch);
У части Hemimetabola (тараканы, богомолы, привиденьевые, клопы) частичное взаимное налегание сложенных крыльев, всегда совершающееся в одном и том же порядке, вызывает диссимметрию крыльев; у кузнечиков и сверчков эта же диссимметрия передних крыльев усилилась благодаря развитию за счет крыльев голосового аппарата. Можно было бы привести ряд других примеров, но и этого достаточно чтобы видеть, что диссимметрия встречается у насекомых в виде множества разрозненных проявлений, но не отражается сколько-нибудь глубоко на общей организации животного и возникновение ее не создает сколько-нибудь значительных рядов развития. У ракообразных мы также встречаемся с многочисленными и разнообразными проявлениями диссимметрии. У Ostracoda наблюдаются довольно часто незначительные проявления асимметрии створок раковины. У Copepoda диссимметрия чаще всего охватывает конечности самца, измененные в качестве совокупительных придатков (последняя пара ног) и органов придерживания самки (антенны); нередко те же конечности развиты диссимметрично и у самок, а иногда эта диссимметрия проявляется уже у неполовозрелых особей. Иногда в корреляции с диссимметрией половых конечностей развиваются асимметрические выросты на боках последнего сегмента груди и первого сегмента брюшка или диссимметрическая форма хвостовой вилки; очевидно, это должно компенсировать при плавании нарушения прямолинейного движения, вызванные диссимметрией конечностей. У Botryllophilus (Copepoda), паразитирующего в жаберной полости асцидий, диссимметрично развиты торакальные ноги: на одной стороне тела они имеют строение плавательных ног, на другой стороне – крючковидное строение. Эта диссимметрия, очевидно, вызвана приспособлением к жизни в узких пространствах полостей хозяина. Резкую диссимметрию всего тела мы наблюдаем у Phryxus (семейство Bopyridae) и других паразитических Isopoda, живущих в жаберных камерах десятиногих раков; в этом случае диссимметрия паразита также обусловлена необходимостью применяться к асимметрической форме тесной полости, в которой он живет (рис. 62, Г). Аналогичное происхождение приписывается обыкновенно диссимметрии брюшка раков-отшельников (Decapoda Paguroidea), прячущих брюшко в раковины брюхоногих моллюсков. Среди отшельников представители семейств Pylochelidae и Pomatochelidae имеют первично-симметричное брюшко, несущее нормально развитые придатки, и прячут его в столь же симметричные раковины Dentalium (Scaphopoda) или в утонувшие кусочки бамбука (рис. 63, А). Представители семейства Paguridae обладают мягким брюшком, которое они прячут в трохоидно завернутых раковинках брюхоногих и которое, в соответствии с этим, асимметрически искривлено и имеет асимметрически развитые тергиты (рис. 63, В);плеоподы 2–5 правой стороны у самок отсутствуют (у самца они отсутствуют с обеих сторон). Coenobita – прибрежные и береговые отшельники Индийского и Тихого океанов – имеют такое же диссимметричное брюшко, как и Paguridae; плеоподы самки, помимо 6-го сегмента, имеются только на 2 и 3-м сегментах правой стороны. Наряду с этими формами, прячущими брюшко в трубку, существуют параллельные им группы, ведущие свободный образ жизни и подгибающие брюшко под гнатоторакс, наподобие крабов. Среди них семейство Lomisidae имеет вполне симметричное брюшко и явно связано с Pomatochelidae. Род Birgus (пальмовый вор) (рис. 63, Г),относящийся к семейству Coenobitidae, имеет почти симметричное строение брюшка, но асимметрия плеопод выдает его родство с ценобитами. Birgus, пройдя личиночное развитие (стадии zoea и leucothoe), покидает море. В литоральной зоне личинка надевает раковинку на еще симметричное брюшко и выходит на берег. После нескольких линек брюшко становится резко диссимметричным; одновременно развиваются легкие, а органы чувств перестраиваются на воздушный лад. Личинка живет в раковине до тех пор, пока находятся раковины достаточной величины. А когда поместительных раковин больше не оказывается, приходится обходиться без них. Время освобождения от раковины зависит от местной фауны наземных моллюсков и в соответствии с этим колеблется в пределах от 6 до 14 месяцев. После освобождения брюшко подгибается на брюшную сторону тела и становится внешне симметричным. На раковинной стадии своей жизни Birgus не отличается от типичных ценобит. Сем. Lithodidae своим коротким панцирем и подогнутым брюшком имитирует крабов (рис. 63, В).Однако тергиты брюшка резко асимметричны, и расположение их указывает на родство литодид с Paguridae; как и у этих последних, самки литодид имеют 3 левые плеоподы. С исторической и экологической точек зрения эти факты из сравнительной анатомии Paguroidea естественней всего объясняются предположением, что диссимметрия брюшка возникла у Paguridae и Coenobitidae вследствие приспособления к жизни в трохоидно-завитых раковинах улиток. Сохранение диссимметрии у Birgus, ведущего во взрослом виде свободный образ жизни, легко объясняется жизнью личинки в трохоидно-завитых раковинах. Для Lithodidae приходится принять, что их диссимметрия унаследована от предков, живших в раковинах. Другой широко распространенный случай диссимметрии у десятиногих раков представляет гетерохелия, т.е. различная форма и величина обеих клешней на первой паре грудных ног (рис. 62, Д). У примитивных, гомойохольных форм обе клешни являются хватательными, у гетерохельных одна из них принимает форму так называемой дробящей клешни и более крупные размеры. Дробящей может быть или правая клешня или левая, или же особи с правыми и левыми дробящими клешнями встречаются в более или менее одинаковом числе. Гетерохелия встречается во всех группах десятиногих, кроме креветок (Natantia). Действительно, асимметрия клешней может развиваться только у донных видов, которые не нуждаются в точном балансировании при плавании. У тех из крабов, которые вторично перешли к плавающему образу жизни, гетерохелия сохраняется в форме клешней, но размеры их выравниваются. Гетерохелия встречается во всех семействах ползающих раков но наряду с этим в большинстве семейств есть и гомойохельные формы. Палеон-тологически гомойохелия древнее; для каждой современной гетерохельной формы, у которой нет современных гомойохельных родственников, они находятся в ископаемом состоянии (Homarus – Eryma -j-, Nica – Blaculla -j-).
Рис. 63. Диссимметрия брюшка у раков-отшельников (Anomura Paguroidea) А – Pylodielea priersii – симметричен, живет в утонувших кусочках бамбука и т.п. (по Smith) Б – Pagurus bernhardus – асимметричен, живет в раковинах брюхоногих (по Cuvier); В – Lithodes maja, самка с брюшной стороны – асимметричная, свободношивущая; 1и 5 – латеральные склериты III и V сегментов брюшка; 2 – маргинальные складки; 3 – последняя пара плеопод; 4 – тельсон и 6-й сегмент брюшка (по Smith); Г – Birgus latro, пальмовый вор, самка с брюшной стороны – наземный, свободноживущий асимметричный вид (по Smith)
Почти в каждом из семейств, известных с мезозоя, мезозойские представители являются гомойохельными, затем в конце мезозоя или в третичную эпоху появляются гетерохельныо формы; отдельные роды почти в каждом семействе эпистатически[5] остаются гомойохельными до наших дней. Гетерохелия в большинстве рядов исторически появляется раньше у самцов, позже – у самок. В настоящее время у ряда форм гетерохельны оба пола. У обычно гомойохельных самок рода Xantho мужской признак дробящей клешни встречается изредка при нормальном развитии женских половых органов. Таким образом, среди десятиногих раков мы встречаемся со значительным развитием диссимметрии в двух различных частях тела – в клешнях многих форм и в брюшке – у части Anomura.
Поиск по сайту: |