|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Метамерия олигомерных аннелид и моллюсков
Олигомерные аннелиды все относятся к полихетам, не представляя в пределах этого класса какой-либо однородной группировки. Однако в отношении характера своей метамерии все лучше изученные олигомерные аннелиды достаточно сходны между собой. Кратко рассмотрим две важнейшие относящиеся сюда группы: семейство Dinophilidae из отряда Eunicemorpha и отряд Myzostomida. Тело Dinophilus (рис. 28) несет несколько венчиков ресниц: два пре-оральных на головной лопасти и пять, расположенных метамерно на туловище. Нервный аппарат состоит из головного мозга, окологлоточных коннективов и двух широко расставленных брюшных стволов, которые несут по 5 ганглиев, попарно соединенных между собой пятью поперечными комиссурами. Выделительный аппарат состоит из 4 или 5 (смотря по виду и полу) пар протонефридиев. Целом представлен гоноцелем (половой полостью), расположенным в задней части тела, и открывается наружу парой выводных протоков (целомодуктов).
Таким образом, у этого животного метамерия простирается на ресничный покров, на нервный аппарат и на протонефридии, т.е. исключительно на органы эктодермального происхождения. Метамерия всех этих органов согласована: имеется одинаковое число метамер кожно-ресничных, нервных и выделительных. Кишечник, мускулатура, половой аппарат неметамерны. Метамерия эктодермальных органов Dinophilus чисто анатомически представляется метамерией упорядочения, точь-в-точь такой же, какую мы видели и у низших червей, например у Procerodes lobata. Точно так же и с эмбриологической точки зрения никаких возражений против подобного толкования метамерии Dinophilus не существует: ларвальные сегменты, имеющиеся у Dinophilus во всех случаях, где их развитие известно, возникают на протяжении тела личинки большей частью все одновременно, без какого-либо процесса, напоминающего подавленное поперечное деление. Таким образом, метамерия Dinophilus возникает в результате метамерного упорядочения в расположении полосок ресничного эпителия, поперечных комиссур нервного аппарата и протонефридиев. Единая метамерия всех этих трех типов образований, т.е. равенство числа нефридиев, венчиков ресниц и комиссур, является, вероятно, также результатом вторичного упорядочения. Среди моллюсков склонность к метамерии выражена сравнительно слабо, наиболее яркое исключение составляют хитоны. Но и на немногие имеющиеся ее проявления исследователи обращали мало внимания, так как поиски метамерии у моллюсков обычно шли по ложному следу. Чаще всего пытались обнаружить у них какие-либо следы целомической метамерии, подобной таковой полимерных аннелид, существование которой у них сомнительно. Вместо этого мы видим у ряда моллюсков проявление той же метамерии упорядочения органов, главным образом эктодермальных, которую мы видели у олигомерных аннелид. Хитоны (Amphineura, Loricata) обладают метамерией, весьма сходной с метамерией олигомерных аннелид: она также охватывает в основном эктодермальные производные и является метамерией упорядочения. В частности, у хитонов метамерной является прежде всего раковина, состоящая из 8 пластинок, из которых 7 лежат на спинной стороне туловища и 1 на спинной стороне головы. Все они располагаются в ряд, вдоль продольной оси взрослого животного (рис. 29, А).В связи с метамерией раковины стоит и метамерия двигающей ее мускулатуры и частичная метамерия артериальной системы. Кроме того, к каждой из пластинок раковины прикрепляются два пучка дорсовентральных ретракторов ноги, которые, в силу этого, располагаются метамерными парами. Жабры (ктенидии) хитонов сидят метамерно в два ряда, по одному ряду с каждой стороны ноги, под краем мантии (рис. 29, Б).Однако метамерия их не согласована с метамерией раковины, так как число жабр гораздо больше, чем число пластинок раковины. Кроме того, у некоторых хитонов происходит укорочение рядов ктенидиев, что делает бранхиомерию тела еще менее совершенной. Метамерия нервного аппарата хитонов слабо выражена, так как поперечные комиссуры между стволами очень многочисленны и расположены довольно неправильно. Но нервы, отходящие от плевровисцеральных стволов к жабрам, строго метамерны. Что касается соленогастров (Solenogastres), раковина у них отсутствует (если не считать диффузно разбросанных спикул), жабры, если имеются, то в числе не более одной пары. Зато поперечные комиссуры между обоими педальными стволами, а также между педальным и плевровисцеральным стволом каждой стороны приобретают у них необыкновенно правильное метамерное расположение (рис. 30, А). Метамерия соленогастров – это почти исключительно метамерия центрального отдела нервного аппарата. Никаких проявлений метамерии целома ни у кого из соленогастров нет, но печеночные отростки отходят иногда метамерно, напоминая этим метамерные ветви кишечника Protomyzostomum. Итак, метамерия в подтипе Атphineura, у наиболее примитивных из моллюсков, в основном сходна с метамерией олигомерных аннелид. По существу – это метамерия упорядочения, далеко не очень совершенная и неполная, охватывающая немногие отдельные аппараты и системы органов, главным образом эктодермального происхождения, и притом не всегда одни и те же у близких между собой форм. Она носит примитивный характер, несовершенный и неустановившийся. Если сравнить эту метамерию с метамерией турбеллярий, например Procerodes, мы обнаружим большое сходство: в обоих случаях имеет место метамерия упорядочения. Но разница еще больше, чем сходство, и обусловлена принципиальным различием планов строения плоских червей и трохофорных животных.
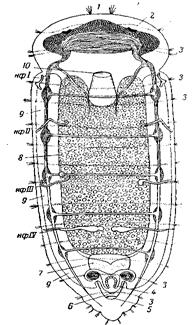
Рис. 29. Метамерия моллюсков А – Tontcia fastigiata (Loricata), молодая особь со спинной стороны; метамерия раковины и раковинных глаз (по Pelseneer); Б – Boreochiton cinereus (Loricata), с брюшной стороны, метамерное расположение ктенидиев (по Pelseneer); 1 – рот; 2 – щупальца; 3 – жабры; 4 – анус; 5 – край мантии; 6 – верхнее отверстие раковины, ведущее в мантийную полость; 7 – раковина; 8 – голова; 9 – эпиподий; 10 – нога; 11 – правый глаз; 12 – гермафродитное половое отверстие; 13 – жаберные придатки; 14 – ринофоры (задняя пара головных щупалец); 15 – головные щупальца
У всех плоских червей продольной осью тела является первичная главная ось, идущая от анимального полюса к вегетативному; эта же ось является у Procerodes и осью метамерии: вдоль нее располагаются все метамерно повторяющиеся органы его тела. Совершенно иное мы видим у хитона или Dinophilus: продольная ось их тела на большей части своей длины параллельна оральной поверхности тела. Следовательно, она перпендикулярна к первичной главной оси посторального отдела, хотя и лежит, благодаря регуляции осей, на геометрическом продолжении главной оси верхнего полушария. Таким образом, ось метамерии аннелид и моллюсков перпендикулярна к оси метамерии турбеллярий. Наглядней всего вытекающие отсюда различия видны на метамерии нервного аппарата; у турбеллярий метамеризуются комиссуры между меридиональными нервными стволами, у трохофорных животных – комиссуры циркумбластопорального сплетения.
Наряду с чисто внешним сходством метамерии обеих групп животных, по существу, между ними имеется принципиальное различие, вытекающее из различий основного плана строения тех и других. У примитивных форм Conchifera склонность к метамерному повторению частей проявляется более или менее в тех же формах, как и у Amphineura. Однако образование на спине единой раковины ведет к тому, что проявления метамерии почти ограничиваются у них мантийной полостью и вентральной (бластопоральной) стороной тела. При наличии очень высокого внутренностного мешка, одетого в длинную трубчатую раковину, прямую или спиральную, нередко возникает частичная метамерия раковины. Особенно хорошо она выражена в раковинах четырехжаберных головоногих с их многочисленными, метамерно расположенными перегородками. При этом осью метамерии является ось внутренностного мешка, не являющаяся одной из осей тела моллюска. В этом случае мы имеем перед собой не метамерию животного в целом, а метамерное расчленение одного из отделов или придатков тела, явление, с которым мы встречаемся и в других группах (например, метамерия рук у офиур или метамерия конечностей у членистоногих). Как существенный общий результат рассмотрения явлений метамерии у олигомерных аннелид и моллюсков мы должны отметить у них наличие наклонности к метамерному расположению органов, главным образом эктодермальных, вдоль оси, параллельной оральной стороне животного, при полном отсутствии метамерии в строении целома.
Вопросы для самоконтроля
1. Метамерия как особый вид симметрии. 2. Метамерия у простейших. 3. Метамерия у колоний кишечнополостных. 4. Метамерия турбеллярий. 5. Классификация строения ленточных червей по Фурману. 6. Теории метамерии цестод. 7. Особенности метамерии олигомерных аннелид. 8. Метамерия моллюсков.
ТЕМА 5
Поиск по сайту: |