|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Архитектоника паразитических плоских червей
Симметрия ленточных червей довольно сложна и разнообразна. У членистых форм сколекс благодаря расположению своих ботридий, хоботков и прочих прикрепительных органов обладает либо 4-лучевой симметрией (Tetraphyllidea – рис. 19, A; Tetrarhynchidea), либо 2-лучевой (Diphyllidea, многие Pseudophyllidea, Cyclophyllidea, рис.19, Би В); реже (у специализованных форм) он принимает какую-либо иную форму. Нервный аппарат сколекса (рис. 18, Б), даже 4-лучевого, всегда обладает 2-лучевой симметрией. Стробила бывает сплющена дорсовентрально, но спинная и брюшная стороны проглоттид мало различаются между собой, в силу чего можно было бы говорить о 2-лучевой симметрии стробилы. Однако здесь 2-лучевая симметрия нарушается строением полового аппарата и расположением полового отверстия (или половых отверстий); последнее лежит чаще всего на брюшной стороне или на одном из боковых краев проглоттиды. В силу этого проглоттиды в первом случае приобретают двустороннюю симметрию, во втором становятся даже диссимметричными. Во внутреннем строении наиболее интересна симметрия нервного аппарата (рис. 18). В туловище многих цестод мы находим две пары спинных продольных стволов, одну пару боковых и две пары брюшных, т.е. те же пять пар стволов и в том же расположении, как у наиболее типичных из бескишечных турбеллярий. Другими словами, мы наблюдаем здесь сочетание 2-лучевой симметрии с 8-лучевой. Иногда встречается и 12 продольных стволов, что изредка наблюдается и у Turbellaria Acoela (удвоение боковых нервных стволов). Наряду с этим у некоторых других цестод (например, Moniezia из отряда Cyclophyllidea) встречается и Значительное развитие лучевой симметрии у ленточных червей исторически является отчасти вторичным, будучи связано с их эндопаразитическим образом жизни и отсутствием подвижности. Однако такое усиление лучевой симметрии могло произойти у цестод лишь благодаря наличию благоприятной почвы – слабо еще закрепленной двусторонней симметрии турбеллярий. Утрата почти всеми ленточными червями способности к правильному поступательному движению ставит перед нами еще один вопрос. Из-за отсутствия поступательного движения цестоды лишены переднего и заднего концов в физиологическом смысле этого слова. Относительно них мы можем говорить лишь о сравнительно-анатомически переднем и заднем концах тела. Другими словами, возникает вопрос: который из концов тела ленточных червей гомологичен переднему концу тела турбеллярий. На этот счет до последнего времени существовали два взгляда: классическая теория, поддерживаемая большинством авторов, считает сколекс передним концом, R. Moniez считал его задним концом.
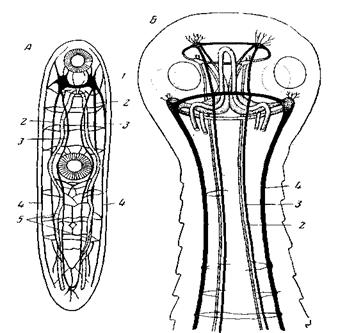
Рис. 18. Симметрия нервного аппарата паразитических платод А – Distomum isostomum (Dlgenea), вид со спинной стороны (из Hanstrom); 1 – мозг; продольные нервные стволы: 2 – брюшные; 3 – спинные; 4 – боковые; 5 – ветви кишечника
Рис. 19. Форма симметрии сколекса у ленточных червей А –4-лучевая симметрия Echinobotlirium flexile (Tetraphylliclea) (из Fuhrmann); Какие же данные существуют за или против гомологии сколекса переднему концу тела турбеллярий? В пользу наличия такой гомологии многие авторы приводили мощное развитие нервного аппарата сколекса, приравнивая его мозгу турбеллярий. Однако многие Monogenea обладают и на заднем конце тела сильно развитым прикрепительным аппаратом, нервный аппарат которого превосходит по своему объему мозг этих форм. Поэтому, если мы представим себе, что сколекс является не передним, а задним концом тела, то его сложный нервный аппарат легко признать новообразованием, вызванным возникновением мощного прикрепительного аппарата. При отсутствии или слабом развитии последнего, как, например, у Аmphilina, центральный нервный аппарат развит на обоих концах тела более или менее одинаково. Таким образом, на основании строения нервного аппарата сказать, который из концов тела является у цестод передним, всегда затруднительно. Внутри класса цестод гомологии концов тела довольно ясны благодаря наличию у личинок всех цестод одинаково расположенных крючков. У метамерных форм крючки всегда располагаются на конце тела, противоположном месту образования сколекса. Расположение крючков у личинок Cestodaria показывает, что у Amphilina гомологичным сколексовому является тот конец тела, который несет хоботок, у Gyrocotyle – тот, который несет присоску. Личинки цестодарий, Amphilina и Gyrocotyle, так называемые ликофоры (рис. 20, Б),несут на одном конце тела по десять крючьев, на другом (сколексовом) – скопление одноклеточных желез. Решающим является сравнение цестод с Monogenea, проведенное Выделительный аппарат большинства цестод очень напоминает таковой прямокишечных, причем изгиб главных стволов, у всех Rhabdocoela приходящийся на область мозга, у цестод имеет место в сколексе.
А – Gyrocotyle Umbriata (отр. Gyrocotyloidea, подкласс Cestodaria), организация (по Lynch, из Hyman): 1 – присоска; 2 – отверстие вагины; 3 –вагина; 4 – семенники; 5 – конечный мешок матки; 6 – желточники; 7 – матка; 8 – семяприемник; 9 – гермарий; 10 – ганглии розетки; 11 – складки розетки; 12 – нервное кольцо розетки;
Таким образом, передним концом цестод несомненно является их сколексовый конец, и в этом отношении права классическая теория. В силу этого все цестоды, развивающиеся при помощи цистицеркоида или цистицерка, с момента метаморфоза и отпадения церкомера лишаются своего заднего конца. То же происходит с другими цестодами с момента отпадения первой зрелой проглоттиды.
а – развитие с корацидием; б – развитие без корацидия; 1 – Pseudophyllidea; 2 – Tetraphyllidea (Proteocephalldae); 3–5 –Cyclophyllidea
Поиск по сайту: |