|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Архитектоника круглых червей
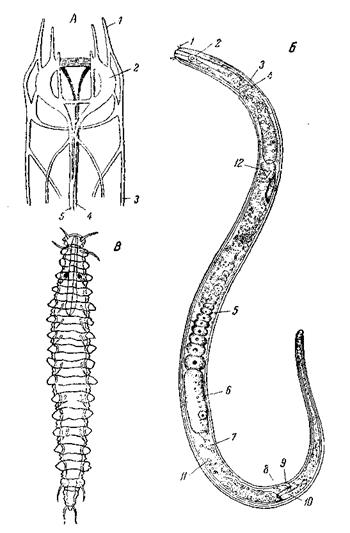
Среди многочисленных групп червей, относимых к Nemathelmintes: Gastrotricha, Rotatoria, Nematodes, Kinorhyncha, Nematomorpha, Priapu-loidea, Acanthocephala, лишь первые три поддаются более или менее точному морфологическому анализу и сравнению с плоскими червями. Относительно всех остальных приходится судить лишь по аналогии, в меру их сходства с первыми тремя группами. Существенными отличиями нематод, гастротрих и коловраток от плоских червей являются наличие заднего прохода. Задний проход Nemathelmintes представляет такое же вторичное сообщение кишечника с внешней средой, как кишечные поры, встречающиеся у некоторых плоских червей. По крайней мере, ни развитие его, ни строение не дают повода видеть в нем что-либо большее или принимать его за гомолог ануса аннелид. Нет оснований и к тому, чтобы считать анальные отверстия в разных классах немательминтов строго гомологичными между собой. Появление их, являясь крупным органологическим усовершенствованием кишечного аппарата, мало отражается на общем плане строения этих животных. Анатомическое сравнение немательминтов не оставляет никакого сомнения в том, что эти животные тесно связаны с плоскими червями. Черты этого сходства распределены по всем трем основным классам немательминтов, причем каждый из них сохранил иные черты. Начнем с нематод. Вытянутое тело нематоды (рис. 22) несет на переднем конце рот; анальное отверстие лежит на брюшной стороне. Вблизи рта лежит глотка, напоминающая pharynx bulbosus турбеллярий и неправильно обозначаемая нематодологами как пищевод. Нефридиев нет. Кожно-мышечный мешок представлен только продольной мускулатурой. Мерцательного эпителия нет. Число клеточных элементов необычайно мало и в основном постоянно. Вся эта организация настолько упрощена и обеднена, что для каких-либо убедительных сравнений как будто не дает материала. Нервный аппарат нематод в основных чертах повторяет расчленение и своеобразную симметрию нервного аппарата плоских червей. Действительно, нервный аппарат всех нематод, так же как и у плоских червей, представляет систему меридианально расположенных продольных стволов, соединенных между собой кольцевыми комиссурами и расположенных радиально вокруг продольной оси тела (рис. 22, А). С первого взгляда между нервными аппаратами обеих групп имеется большое различие: для нематод весьма характерны два непарных нервных ствола – спинной и брюшной, плоские черви непарных стволов никогда не имеют. Спинной, и брюшной стволы Oxyuris начинаются от мозговой комиссуры, каждый двумя корешками; на заднем конце тела оба ствола также расщепляются. Встречая на своем пути женское половое отверстие, брюшной ствол расщепляется и здесь, кольцом охватывая вагину. Наконец, в нескольких случаях у нематод были описаны и парные брюшные стволы, а именно – у молодой Toxocara mystax, у Strongylus convolutus и у Plexus. Все это с несомненностью говорит о парном происхождении обоих стволов. Если же допустить, что брюшной ствол Oxyuris соответствует паре брюшных стволов и спинной – паре спинных, то в таком случае мы возвращаемся к наиболее распространенному среди платод типу нервного аппарата с тремя парами продольных стволов. Передние головные нервы большинства нематод отходят тремя радиально расположенными парами, что нередко связано со столь же радиально-симметричным расположением головных органов чувств. Мало того, у свободноживущих морских нематод встречается и большее число продольных стволов, до 5 или 6 пар, что соответствует максимальному числу нервных стволов у низших турбеллярий и цестод. Наличие нескольких пар латеральных стволов и парная природа брюшного ствола были в дальнейшем констатированы у ряда нематод разными авторами. Таким образом, симметрия нервного аппарата нематод совпадает с симметрией его у плоских червей.
А – нервный аппарат Oxyuris cunula: 1– передние нервы; 2 – боковые ганглии; нервные стволы: 3 – боковые; 4 – брюшной; 6 – спинной (по Martini); Б – типичная свободная нематода Monhystera subflliformis, самка справа: 1 – головные щетинки; 2 – амфиды (боковые чувствительные ямки головы); 3 – шейная пора; 4 – нервное кольцо;
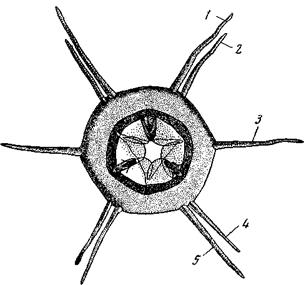
В обоих случаях, помимо двусторонней симметрии, мы находим сочетание неполной 2-лучевой симметрии с неполной 4-лучевой или Помимо центрального нервного аппарата, по этому же типу симметрии располагаются нередко и головные органы чувств нематод
Рис. 23. Голова свободной морской нематоды Cobbia triodonta, Виден трехгранный просвет глотки, три челюсти – спинная и две вентро- латеральные, головной венчик, состоящий из щетинок, расположенных по правилам ктенофорной симметрии: 1 – субдорсальные щетинки; 2 – дорсо-латеральные; 3 – латеральные;
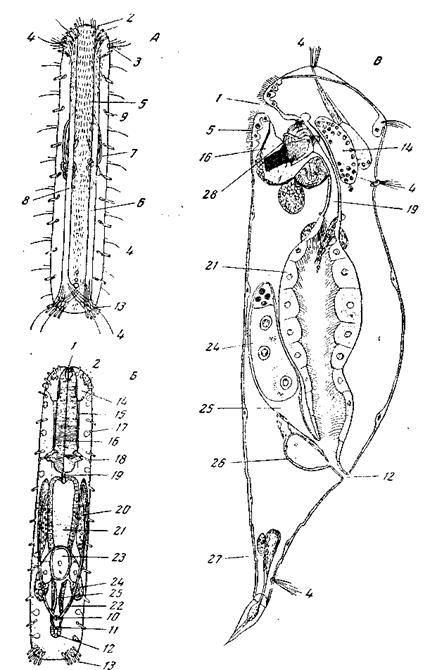
рот ее сдвинут на передний конец тела и кишечный рот, т.е. отверстие, соединяющее глотку со средней кишкой, обычно, хотя не всегда, – на передний конец кишечника, как мы это видим у многих турбеллярий (например, у Prorhynchus из отряда Alloeocoela) и у трематод. Более примитивное положение кишечного рта, на вентральной стороне кишечника, наблюдается у немногих нематод, например, у Роrrосоесит (из отряда Ascaridata). На заднем конце кишечника нематод образовалось анальное отверстие. Все остальные различия по сравнению с турбелляриями сводятся к гистологическим и органологическим изменениям, не отражающимся на основном плане строения. В органологическом отношении между нематодами и турбелляриями существует переходная группа в лице гастротрих. Мы видели, что у плоских червей лучевая симметрия искажается и постепенно стирается из-за все возрастающей роли двусторонней симметрии. У нематод двусторонняя симметрия резко выражена в положении заднего прохода и половых отверстий и отражается также на строении нервного аппарата. Строение гонад большей частью даже диссимметрично. Однако наряду с этим у нематод развивается вторичная радиальная симметрия 4-лучевого типа, выражающаяся в расположении утолщений гиподермы (спинная, брюшная и боковые линии), мышечных полей и т.п. Вторичая радиальная симметрия связана, вероятно, с роющим образом жизни большинства свободноживущих нематод, и ей не препятствует эндопаразитический образ жизни остальных. Эта 4-лучевая симметрия является, конечно, неполной, налагаясь на первичную радиальную симметрию нервного аппарата и на билатеральную основу всей организации. В строении нервного аппарата она проявляется в том слиянии спинных и брюшных стволов в непарные медиальные стволы, которое так затрудняет правильное понимание симметрии нематод. Гастротрихи (Gastrotricha; рис. 24, А, Б)во многих отношениях очень близки к нематодам. Их сближает строение кутикулы ее придатков, строение гиподермы, положение рта, строение глотки, наличие анального отверстия. С турбелляриями их сближает общая форма тела (у многих Масrоdasyoidea), остатки ресничного покрова, плавание при помощи ресниц, наличие протонефридиев, внутриклеточное пищеварение, гермафродитизм (ivlacrodasyoidea), органы чувств в виде мерцательных головных ямок. Гомологами последних являются, вероятно, и амфиды (чувствительные ямки) нематод. Глотка гастротрих, как и глотка нематод, принципиально не отличается от массивной глотки (pharynx bulbosus) плоских червей. Существенными отличиями от турбеллярий являются присутствие кутикулы, дифференцированная туловищная мускулатура и отсутствие кожно-мышечного мешка. Половой аппарат гастротрих по своим особенностям не выходит за пределы его многообразия у турбеллярий. Строение гонад сближает гастротрих с Macrostomida. Мозг лежит дорсально от кишечника и непосредственно примыкает к эпидермису головы, этим напоминая мозг многих мелких турбеллярий. От мозга отходят кзади два продольных нервных ствола, как у Macrostomum. Таким образом, нервный аппарат построен строго билатерально и больше напоминает таковой Macrostomida, нежели нематод. Мы видели, что развитие гастротрих также сближает их с Turbel-laria: ранние стадии дробления напоминают дробление Macrostomum, дальнейший ход развития говорит в пользу гомологии продольной оси тела гастротрих с первичной осью зародыша, а следовательно и с продольной осью тела турбеллярии. В то же время организация взрослых гастротрих для установления гомологии продольной оси тела убедительного материала не дает, т.к. их нервный аппарат утратил радиальное строение. Наоборот, развитие нематод ничего не дает для проморфологи-ческого сравнения их с турбелляриями; зато строение взрослого животного, и в особенности архитектоника нервного аппарата, ясно говорят о единстве плана строения нематод и турбеллярии. Вряд ли может быть сомнение во взаимной близости гастротрих и нематод и, в силу этого, их сходства с турбелляриями взаимно допол-няют друг друга: развитие гастротрих и архитектоника взрослых нематод свидетельствуют о единстве плана строения немательминтов и турбеллярии, строение взрослых гастротрих и развитие нематод не противоречат этому заключению. Понимание истинного прототипа коловраток (Rotatoria) (рис. 24, В)долгое время затруднялось господством трохофорной теории. В применении к коловраткам эта теория принимала двоякий вид. Одни авторы считали коловраток видоизмененными потомками трохозоона, гипотетического предка всех трохофорных животных. Другие авторы считали коловраток неотеническими потомками кольчатых червей. Но и те и другие пытались свести организацию коловраток к организации трохофоры и потому за наиболее типичных коловраток принимали тех из них, которые имеют дифференцированный коловращательный аппарат в виде двух венчиков ресниц (trochus и cingulum) и лишены ноги. Большой поддержкой этой теории явилось в глазах ее приверженцев открытие Земпером аберрантной коловратки Trochosphaera aequatorialis, представляющей грубое внешнее сходство с трохофорой аннелид. Однако и в те времена В дальнейшем обширные исследования, с несомненностью показали, что сравнительная анатомия внутри класса Rotatoria решительно говорит против сближения коловраток с трохофорными личинками, а следовательно, и против приложимости к ним трохофорной теории в обоих ее видах. Развитие коловраток также не представляет ни малейшего сходства с развитием трохофоры. Коловратки не имеют ничего общего ни с трохофорными животными, ни с их личинками. Наиболее примитивными среди коловраток являются донные, ползающие формы из семейства Notommatidae, вроде Proales или Diglena. Они имеют продолговатое тело, мало отличное по форме тела от мелких турбеллярии и особенно гастротрих. Разделение на голову, туловище и ногу едва намечается. Задний конец уже раздвоен, как, впрочем, и у многих Gastrotricha. Мерцательный аппарат представляет поле ресничек, занимающее переднюю часть брюшной стороны животного. Точно такое же строение брюшного мерцательного покрова мы находим а у некоторых гастротрих, как, например, у Hemidasys. Примитивность ползающих нотомматид сказывается, между прочим, и в том, что они обладают сравнительно мало редуцированными самцами. С переходом к плавающему образу жизни в организации коловраток происходит несколько перемен: тело все более резко дифференцируется на голову, туловище и хвост (нога), мерцательный аппарат, постепенно усложняясь, принимает сложно расчлененную форму, происходит прогрессивная редукция организации самцов. У некоторых крайних форм из числа планктических коловраток редуцируется и хвост.
Gastrotricha Macrodasyoidea, схема строения: А – вид со спинной стороны; Б – вид с брюшной стороны (по Remane); В – схема строения коловратки (по Remane) Возникающие в разных семействах типы коловращательного аппарата весьма разнообразны, и коловращательный аппарат в виде двух венчиков ресниц, на который опиралась трохофорная теория, представляет лишь один из многих производных типов этого аппарата. Итак, для сравнения коловраток с другими группами мы можем исходить только из организации примитивных коловраток (сем. Notommatidae). Ничтожные размеры, форма тела, и, в частности, форма его заднего конца, кутикула, характер мерцательного покрова, дифференцированная мускулатура при отсутствии общего кожно-мышечного мешка оказываются общими признаками низших коловраток с гастротрихами. С нематодами их сближает постоянство клеточных элементов. Примитивной чертой, сближающей коловраток непосредственно с турбелляриями, является прежде всего строение протонефридиев; в этом отношении коловратки стоят ближе к турбелляриям, чем к гастротрихам и киноринхам. Далее с турбелляриями их сближает наличие в теле кольцевых мышц, вентральное положение рта. Впрочем, строго терминальный рот гастротрих и нематод при эмбриональном развитии также закладывается на брюшной стороне и лишь постепенно сдвигается на передний конец тела. Своеобразный признак коловраток – наличие кутикулярного вооружения глотки, но среди турбеллярий представители отряда Gnathosto-mulida также имеют глотку, вооруженную кутикулярными челюстями, правда, несравненно более простыми, чем у коловраток, так что способность к образованию глоточного вооружения не чужда, очевидно, и турбелляриям.
Вопросы для самоконтроля 1. Теории происхождения турбеллярий. 2. Особенности строения ленточных червей. 3. Симметрия ленточных червей. 4. Архитектоника нервного аппарата нематод и вопросы происхождения немательминтов. 5. Строение гастротрих и их происхождение. 6. Строение коловраток и их происхождение.
ТЕМА 4 МЕТАМЕРИЯ НИЗШИХ БЕСПОЗВОНОЧНЫХ, АННЕЛИД
Поиск по сайту: |