|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Различия между молодыми и взрослыми особями
Молодые птицы боятся быстро летать. Для них это очень характерно. Вы когда-нибудь вели машину со всей скоростью, на какую она способна? Скорость убивает! Каждый год мы наблюдаем за облетываемыми соколами развивающими свое летное искусство, и совершенно отчетливо видим, что им не нравится быстро летать, потому что тогда они не могут справиться с посадкой. К счастью, на этой стадии их перья еще находятся в пеньках, поэтому при ударах они не наносят себе серьезных увечий. Но если вы посадите молодую птицу, такую как балобан, в вольер и не позволите ей летать до отвердения перьев, то потом, когда он окажется на свободе, он вполне может запаниковать. Он взлетит и разовьет хорошую скорость, прежде чем осознает, что происходит, затем, не умея тормозить, приземляться и даже разворачиваться, может пролететь километр или два, прежде чем упадет. Если он столкнется с деревом и оцарапается о ветки, то потом в течение одного или двух дней может отказываться летать. В это время, сидя под деревом, вы сможете поразмыслить, почему так произошло. Молодые хищники, развивающиеся в естественных условиях, в первые четыре недели учатся маневрировать во время обычного полета. Они играют друг с другом и срывают лиcтья с деревьев, постепенно развивая координацию. При этом родители продолжают обеспечивать их пищей, поэтому они не пытаются воспроизводить опасные маневры охотника за добычей. Именно в этот момент начинает действовать естественны отбор. Один из путей обойти эту проблему - не пытаться поймать быструю проворную добычу. Многие молодые хищники, такие как краснохвоcтые канюки, балобаны и мексиканские соколы в первую осень ведут себя скорее как «ищейки», чем как «нападающие» (см. главу 6). Они ловят легкую добычу типа червяков, насекомых, полевок и других мелких животных. С наступлением зимы большинство таких животных исчезает. Они оказываются запертыми в замерзшей земле, прячутся под снегом или просто вымирают. Молодым хищникам необходимо переключиться на более сложную добычу и стать более «нападающими». Многие этого не делают. Те же, кому это удается, способны к высокоскоростной атаке и могут выжить при меньшем количестве добычи. К следующей весне, когда пищи становится много, они уже утвердятся как охотники и будут способны прокормить не только себя, но и свои семьи. Все это отражается на крыльях. У молодых птиц маховые и рулевые длиннее, чем у взрослых. Это уменьшает нагрузку на крыло и скорость сваливания. Молодые могут летать медленнее, чем их родители, что более характерно «ищейкам». Их перья более мягкие, менее ломкие, но не так эффективны для быстрого полета. У нас много информации по этому вопросу, но она весьма запутана, т.к. разные виды с возрастом изменяются по-разному. У сапсана и дербника, например, взрослые особи имеют более короткие и узкие крылья и более короткие хвосты. У ястребов с возрастом хвосты укорачиваются, но удлиняются и заостряются крылья, они становятся более соколообразны. Чтобы изучить этот вопрос, необходимо 0бобщить информацию относительно оперения молодой и взрослой птицы, для одних и тех же особей, а не отдельную информацию относительно оперения случайных особей. Это одна из перспективных областей для тех, кто ищет тему для исследований.
Планирование и парение
Очень часто можно видеть дикого сокола, который сначала поднимается вверх на раскрытых крыльях с раздвинутым веером хвостом в восходящем воздушном потоке около склона холма, пока не превратится в крупинку в небе, а затем планирует по прямой туда, где собирается охотиться. Используя эти две техники полета, птица с очень небольшими затратами энергии может облететь всю свою территорию. Проведя много времени с соколами, вы сможете «увидеть» эти невидимые подъемы почти так же ясно, как эскалатор в универмаге. Парение является формой планирования, при котором птица все время падает в воздухе. При парении птица необязательно старается далеко пролететь по горизонтaли, а скорее стремится, не меняя местоположения, набрать высоту, чтобы охладиться, выследить добычу или во время токования. При этом акцент делается не на перелет, а на то, чтобы удержаться на высоте. Значит, воздушной скоростью можно пожертвовать, поэтому при данном угле планирования скорость падения птицы будет меньше. Поэтому парители обычно имеют меньший вес. При нормальной скорости полета тормозной эффект пропорционален воздушной скорости, поэтому при меньшей воздушной скорости он менее важен, а значит, птица может иметь большиe «подъемные» поверхности - большие крылья, широкие второстепенные, большой веерообразный хвост. Создаваемый дополнительный тормозной эффект на низких скоростях более чем компенсируется получаемой дополнительной подъемной силой. У парящих видов наружныe первостепенные имеют вырезки, поэтому кончики крыльев напоминают пальцы. Каждый кончик первостепенного действует как отдельная узкая аэродинамическая поверхность, обеспечивая подъем с минимальным тормозным эффектом, а также способствует снижению скорости сваливания. Для достижения этого, первостепенные постепенно становятся жестче по направлению к наружному краю крыла. Крылья срываются с потока, если воздух, вместо того, чтобы свободно просачиваться через крыло, течет через кончик крыла и становится турбулентным. Вырезки перенаправляют воздух через верхушку крыла, уменьшая турбулентность и понижая скорость сваливания, поэтому птица может удерживаться в воздушном потоке при более низкой скорости движения вперед (1.15.4).
Специалисты по парению, сведя свой уровень падения к минимуму, нуждаются в воздушныx потоках, восходящих с большей скоростью, чем скорость их падения. Такие воздушные потоки образуются потоками воздуха, восходящими со склонов холмов при сильном ветре или при достаточном прогреве воздуха, который начинает подниматься от нагретой земли. Очевидно, что в областях с ровным рельефом хищные птицы могут рассчитывать только на восходящие потоки теплого воздуха. В холодном климате на это рассчитывать не приходится, и специалисты по парению, а также некоторые орлы и канюки здесь выжить не могут. Они привязаны к местам, где поднимающиеся воздушные потоки доступны достаточно часто, чтобы можно было найти пищу. Обычно им приходится каждое утро ждать, пока воздух прогреется и начнет подниматься. Фридрих II запрещал своим сокольникам выпускать соколов после 9 часов утра из-за риска нападения орлов, которые в это время начинают парить. Хотя большинство хищников могут планировать и парить, наиболее активные виды хищников, такие как крупные соколы и ястребы, главным образом приспособлены к атакующей фазе охоты (см. 6.1) и способны к мощному машущему полету. В отличие от самолетов, птицы живые и могут в разные моменты изменять свой профиль. Так, сапсан, регулируя форму крыльев и перьев, может оптимизировать свою форму для высокоскоростного планирования, а затем изменить ее для парения. По птицы форме можно понять ее намерения: узкие, немного согнутые крылья и сжатый хвост показывают, что она планирует и собирается пролететь большую дистанцию - вероятно, она увидела вдали какую-то добычу (см. 6.13). Веерообразные крылья и хвост показывают, что птица стремится скорее подняться вверх, чем лететь вперед. Возможно, она нашла восходящий воздушный поток, который поднимет ее до уровня облаков. Виды, имеющие небольшую нагрузку на крыло, такие как канюки, главным образом ищут добычу, которую легко поймать. Они мастера парения и планирования. Они не смогут поймать быструю проворную дичь. Их излюбленный способ нападения планирующая атака (6.13).
Активный полет
Сходным образом летают сарычи и орлы, но взмахи крыльев у них глубже и медленнее, а крылья складываются сильнее. Их крылья движутся вверх быстрее, чем вниз. При напряженном полете весьма вероятно, что взмах становится активен, обеспечивая некоторую подъемную силу, в то время как при более медленном полете, он, вероятно, пассивен. Когда сокол набирает скорость выше 7 метров в секунду, воздушной скорости достаточно, чтобы поднять крылья, что уменьшает нагрузку на надлопаточные мыщцы. Когда взмах активен, он создает следовые завихрения (рисунок 1.16.2). Вместо того чтобы складываться близко к телу, крыло при взмахе удерживается жестче и прямее, что ведет к типичному мерцающему полету крейсирующего сапсана (рисунок 1.16.3). Когда хищник хочет лететь быстрее, чем при крейсирующем полете, у него есть три возможности: сарычи и крупные орлы не могут развить настоящий спринтерский полет, поэтому для более быстрой атаки они должны атаковать сверху на скоростном планировании (см. 6.13). Соколы и ястребы действительно могут лететь быстрее, но делают они это двумя разными способами. Здесь я, похоже, достиг предела того, что на сегоднящний день известно, поэтому могу только поделиться своим мнением по дaнному вопросу. Проблема состоит в том, что с использованием аэродинамической трубы, и без нее, сложно детально изучить хищника во время спринтерского полета. Сокол, который летит на такой большой скоpости, что обычные крейсирующие взмахи крыльев будут только затормаживать полет, похож на велосипедиста, который не может крутить педали достаточно быстро, чтобы ускорить движение. Поэтому сокол скользит в воздухе с полузакрытыми крыльями, при этом второстепенные обеспечивают подъемную силу, в остальном минимизируя рычажную нагрузку на грудные мыщцы. Сзади мягко слетают следовые вихри (в центре, рисунок 1.16.4) как при крутом плaнировании (сравните рисунки 1.15.2 и 3). Затем сокол выполняет серию глубоких пульсирующих взмахов, при которых он вкладывает как можно больше силы в несколько быстрых опусканий крыла, более быстрых, чем скорость полета, и которые направлены таким образом, чтобы обеспечить толчок, а не подъем. Результатом, который очень хорошо заметен у кречета на спринтерском полете, является то, что каждый взмах крыльев явно рывками толкает птицу вперед. Выглядит это так, словно кто-то невидимый пинает птицу сзади. Даже небольшой серии таких пульсирующих взмахов достаточно, чтобы сокол набрал такую скорость, при которой дальнейшие взмахи крыльев бесполезны. К тому времени что-нибудь произойдет.
При опускании крыла (рис. 1.16.5 а-е), создается толчок и подъем, но крыло очень короткое, что дает грудным мышцам техническое преимущество, кончики первостепенных сгибаются назад к тому месту, где угол их наклона достигает 90 градусов к поверхности крыла по вертикали и по горизонтали (рис. 1.16.5 е). Затем птица начинает создавать тягу своими крыльями, активно используя надлопаточныe мышцы (1.16.5 f). Эластичные первостепенные начинают восстанавливать свою нормальную форму, толкая воздух вниз и назад, что обеспечивает подъем и толчки, а также оказывает помощь надлопаточным мышцам. Теперь крыло на полпути назад и наполовину сложено.Края первостепенных направлены вперед и выровнены с углом падения. В верхней точке взмаха они сходятся вместе и снова обеспечивают толчок при опускании крыла.
Обыкновенный сарыч (рисунок 1.16.6) относительно плохо летает активным полeтом. Он медленно разгоняется, имеет небольшую максимальную скорость и вскоре выдыхается. Редко можно увидеть, чтобы сарыч пролетел спритерским полетом более 100 м, очень редко 200 метров. Он скоро начинает отдыхать, планируя между взмахами, на графике это начинается на 40 метрах. При горизонтальном полете он может набрать скорость до 10 мeтpов в секунду, делая около 5-6.5 взмахов в секунду. Немного найдется добычи, которая передвигается достаточно медленно, чтобы сподвигнуть сарыча приложить усилия дольше, чем на 80 метров. Большая часть добычи или будет быстро поймана (например, полевки) или при погоне оставит сарыча Мелкие ястребы, такие как перепелятник, напротив, развивают максимальную скорость менее чем за секунду, на первых же метрах. Их взрывной спринт дает им преимущество над всеми другими перечисленными хищниками. Однако немногие сохраняют спринт на предельной скорости больше чем на 100 м. К этому времени полет обычно так или иначе заканчивается, и если он был успешным, то перепелятник примерно через 150 метров садится. Изучая тетеревятника Slijper обнаружил, что самцы стартуют быстрее, но примерно через 70 метров самки их обгоняют. Встав на крыло, самки летят немного быстрее. Примерно через 130 метров тетеревятники обычно сбрасывают скорость. Если им не удалось поймать жертву в начале спринта, они бросают преследование или летят по инерции, набирая высоту и следя за жертвой. На первых 20 метрах сапсан летит не намного быстрее сарыча, но на 50 метрах он начинает набирать скорость, примерно на 130 метрах он обгоняет тетеревятника и поддерживает хорошую скорость в течение нескольких сотен метров. При горизонтальном машущем полете на длинной дистанции его, вероятно, обгонит только кречет. Новозеландский сокол со своим ястребиным профилем и соколиной физиологией, стартует скорее как небольшой самец тетеревятника. К тому времени, когда самка перепелятника пролетит 80 метров, сокол пролетит 100 метров. Примерно к 130 метрам, когда тетеревятник начинает сбавлять скорость, скорость новозеландского сокола остается прежней, но его обгоняет набравший скорость сапсан. К тому времени, когда сапсан достигнет 280 метров, новозеландский сокол будет отставать примерно на 40 метров, и оба исчезнут за горизонтом без признаков усталости. Ускорение и максимальная скорость у хищников при преследовании должны быть сопоставимы с этими параметрами их жертв. Перепел имеет сходный рисунок полета с перепелятником, фазан с тетеревятником, а голуби (хотя немного быстрее при взлете) как у сапсана. Ястребы быстро бросают сильную, способную к длительному полету добычу, если только не ловят ее на спринте или напав из засады. Эффект взрывного старта ястребов проявляется в наборе скорости в первые 40 метров. Ястреб Купера, например, обычно делает 4-5.5 взмахов в секунду, а при взлете 7-8 взмахов в секунду, используя грудные мышцы, составляющие около 17% от общего веса тела. Он преодолеет эту дистанцию, когда сарыч и большинство соколов пролетят всего 20 метров. Их мастерство в преодолении дистанции состоит в способности выполнить прямую атаку с лета (см. 6.10) и умении оценить максимальную дистанцию, на которой целесообразно атаковать намеченную добычу. Крупные соколы обычно не предпринимают короткие прямые атаки, а предпочитают более предсказуемые длинные. Сарычи по возможности вообще не используют спринтерский полет, вместо этого они используют высоту, позволяющую атаковать с планирующего полета или спикировать. Кроме бесконечных преобразований кривых разгона и дистанций атаки, существует проблема маневрености. Здесь наивысшие показатели у ястребов, дербника и новозеландскаго сокола; крупные соколы и ястребы Харриса менее проворны и более громоздки в хвостовой части, чем сарычи. Добыча тоже очень различается по маневрености (см. раздел 7.4). Обычно ценой большей маневрености является меньшая максимальная скорость, длинный хвост способствует верткости, но создает тормозной эффект.
Строение лап
В основе лап лежит последовательность костей и заключенных в оболочки сухожилий (рисунок 1.17.1). За голенью и цевкой проходят очень сильные сухожилия мышц-сгибателей. Они контролируются мышцами, которые лежат выше голени.Сухожилия мышц-сгибателей уходят прямо вниз через подъем свода стопы нижней части каждого пальца к дальним подушечкам. При сокращении мышц, лапа сильно сжимается. Сухожилия скользят в канавках нижней cтoроны костей пальцев и удерживаются на месте жесткими оболочками. Оболочки имеют тонкие гребешки, напоминающие бороздки на отпечатках пальцев и сцеплены с жесткими рубцaми на сухожилиях (рисунок 1.17.2). Когда лапа сжимается, эти рубцы сцепляются вместе, как храповый механизм ручного тормоза. Возможно, вы замечали жесткий, отрывистый скрип при попытке ослабить хватку хищной птицы. Это происходит при попытке силой пересилить храповик. Храповик выполняет две полезныe для птицы функции, он позволяет спящей птице «запирать» лапы, сжимающие ветку, а также гарантирует, что удержание сильной добычи не потребует больших мышечных усилий. Когда человек пытается крепко схватить что-нибудь рукой, он может удерживать хватку лишь несколько секунд, после чего мышцы начнут терять силу; если жертва замрет, Сухожилия сгибателей идут по бороздкам в верхней части цевки таким образом, что вес спящей птицы приводит в действие храповик, что усиливает хватку за присаду-насест. Однако на плоской поверхности, к примеру, тетеревятник, сухожилия которого настроены на сцепление, чувствует себя некомфортно. Он не может полностью разжать лапы, не выпрямив полностью лапу, но в таком положении ему трудно сидеть. Если его держать в течение нескольких недель на плоской поверхности, его мышцы-сгибатели могут сильно ослабеть. Сухожилия мышц-разгибателей идут вниз в передней части голени и цевки, одно из них ответвляется назад связкой, идущей к заднему пальцу. Их функция - разгибать пальцы, поэтому они слабее сгибателей. Две связки действуют наподобие блоков и удерживают сухожилия на месте, иначе бы они вышли наружу. Кровеносные сосуды лежат сразу под кожей, так что сухожилия не могут их зажать. Большую часть пути артерии и вены идут параллельно. Это значит, что теплая идущая по артериям кровь охлаждается кровью, идущей по венам, а та в свою очередь, нагревается. Это способствует сохранению тепла в холодную породу, а при физических нагрузках или в жаркую погоду сосуды расширяются, позволяя терять большее количество тепла. Некоторые грифы даже мочатся на свои лапы, чтобы увеличить охлаждение посредством испарения. К счастью, ловчие птицы не имеют такой привычки! Поскольку лапы в основном состоят из костей и сухожилий, кровообращение в них довольно скудное, особенно во время долгого бездействия. Это значит, что любое повреждение, растяжение или разрыв связок, укус, прокол шипом или царапина может потребовать длительного времени для заживления. С инфекциями тоже трудно бороться. Длительное напряжение может привести к дегенерации подлежащей ткани или к формированию области омертвевшей ткани, типа мозоли. Нижняя сторона лапы имеет шишковидные отростки. Если птицу держать на гладкой поверхности, давление будет приходиться на них. Сначала они станут гладкими и могут порозоветь, затем они медленно отомрут, открывая ворота инфекции. Окружающая ткань отреагирует созданием нескольких слоев клеток, что приведет к образованию изуродованной стопы.
Профилактические меры позволяют уменьшить подобный риск. Птиц лучше содержать на присадах, имеющих достаточную кривизну, для предотвращения чрезмерного натяжения сухожилий-сгибателей, но в тоже время достаточно широких, чтобы вес птицы распределялся равномерно по всей площади ступни. Рифленая поверхность, типа астротурфа, предотвращает давление на отдельныe точки, не вызывая истирания, а водоотводящие поверхности предупреждают намокание ступней, а также уменьшают вероятность возникновения бактериальных инфекций. Сбалансированное питание с дачей костей и сухожилий стимулирует образование здоровых тканей и хорошее кровоснабжение, предупреждая «опухание» лап и уменьшая вероятность того, что птица будет клевать их от скуки. Вольер должен быть оборудован таким образом, чтобы предотвратить жесткое приземление птицы, могущее повредить лапы. Кормление по утрам и контролирование веса птицы позволяют уменьшить давление на лапы, т.к. днем пища переваривается быстрее (см. раздел 5.9). Аккуратное подрезание когтей, умелое обращение с птицей и немедленное залечивание укусов и проколов уменьшит риск инфицирования. Хотя способы лечения лап постоянно совершенствуются, профилактика все же лучше лечения. Когти различаются по форме и размеру. Задний и внутренний когти служат в основном для удержания и умерщвления добычи, поэтому они самые длинные. Коготь центрального пальца (2 палец) короткий с латеральным выступом с внутренней стоpоны. Хищная птица использует его для чистки и расчесывания нежных участков головы, которые можно повредить при расчесывании острыми когтями. Коготь наружного пальца тоже короткий. Эти два пальца придают хищной птице устойчивость и увеличивают вероятность захвата добычи. Кожа лап здоровых взрослых особей большинства хищных птиц светлоокрашенная благодаря наличию каротина. У крупных соколов взрослые самцы-производители имеют заметно более ярко окрашенные желтые лапы, чем самки, лапы которых бледнее. Возможно это происходит из-за того, что каротин используется при развитии яиц. При ухаживании самцы демонстрируют свои яркоокрашенные лапы, прохаживаясь «на носочках» (рисунок 2.6.2). Пропорции лап варьируют в зависимости от добычи. Хищники, которые ловят добычу в воздухе, например, перепелятник, сапсан, дербник, красногрудый сокол (Falco deiroleucus), новозеландский сокол, имеют длинные пальцы, обеспечивающие быстрый захват. Сапсаны имеют короткую плюсну, способную противостоять ударам при пикировании, а длинная цевка перепелятников и новозеландских соколов позволяет им хватывать птиц в зарослях и ловить их во время маневренной погони. Териофаги, такие как пустельги и мноrие виды канюков, имеют более короткие пальцы, поскольку умерщвление добычи происходит посредством сильного сжатия лапы, а не удара клювом. Виды, охотящиеся и на птиц, и на млекопитающих, такие как тетеревятники, балобаны, имеют промежуточное строение лап, отражающее их двойную направленность. Существуют специализированные виды птиц, например, птица-секретарь, у которой длинные лапы и небольшие сильные пальцы, необходимые для ловли змей. Внимательное обследование лап показывает, что они столь же приспособлены к образу жизни, как и крылья.
Поиск по сайту: |
 Планирование и парение являются формами пассивного полета с фиксированными крыльями, который зависит от трех сил: подъемной силы, веса и сопротивления (рис. 1.15. 1). Птица двигается вперед по воздуху под действием гравитации, снижаясь под углом к земле. Движение вперед генерирует подъем через аэродинамический профиль крыла, но уровень подъема меньше, чем уровень падения. Если скорость подъема воздушной массывыше скорости падения птицы, то последняя сможет набрать высоту; это парение. При планировании, чтобы лететь вперед, птица падает в воздухе, при этом высота переходит в дистанцию, в то время как при парении птица поднимается благодаря воздушным потокам, у которых скорость подъема выше, чем скорость падения птицы. Парение является формой планирования, при котором для набора высоты используется поднимающийся воздух. При планировании птица медленно теряет высоту с увеличением пройденной дистанции. Отношение высоты к пройденной дистанции называется углом планирования. Умеренно способная к планированию птица с длинными узкими крыльями, например, взрослый сапсан, достигает угла планирования 6 или менее градусов при отношении примерно 1 к 10. Это означает, что с высоты в 300 метров (1000 футов) птица в неподвижном воздухе пролетит по горизонтали 3 километра (около 2 миль) или больше. Это достигается уменьшением тормозного эффекта за счет жесткой обтекаемой формы, длинных крыльев и короткого хвоста, а не за счет уменьшения веса. Приспособленность к тому или иному полету изменяет только скорость снижения, но не влияет на угол планирования. Тяжелый сапсан способен достигнуть такого же угла планирования, как и легкий, но при планировании он движется быстрее. Сглаженный контур минимизирует возмущения окружающего воздуха. Воздух гладко огибает крыло и, скручиваясь, срывается со свисающего края кончика крыла наподобие вихря (рисунок 1.15.2).
Планирование и парение являются формами пассивного полета с фиксированными крыльями, который зависит от трех сил: подъемной силы, веса и сопротивления (рис. 1.15. 1). Птица двигается вперед по воздуху под действием гравитации, снижаясь под углом к земле. Движение вперед генерирует подъем через аэродинамический профиль крыла, но уровень подъема меньше, чем уровень падения. Если скорость подъема воздушной массывыше скорости падения птицы, то последняя сможет набрать высоту; это парение. При планировании, чтобы лететь вперед, птица падает в воздухе, при этом высота переходит в дистанцию, в то время как при парении птица поднимается благодаря воздушным потокам, у которых скорость подъема выше, чем скорость падения птицы. Парение является формой планирования, при котором для набора высоты используется поднимающийся воздух. При планировании птица медленно теряет высоту с увеличением пройденной дистанции. Отношение высоты к пройденной дистанции называется углом планирования. Умеренно способная к планированию птица с длинными узкими крыльями, например, взрослый сапсан, достигает угла планирования 6 или менее градусов при отношении примерно 1 к 10. Это означает, что с высоты в 300 метров (1000 футов) птица в неподвижном воздухе пролетит по горизонтали 3 километра (около 2 миль) или больше. Это достигается уменьшением тормозного эффекта за счет жесткой обтекаемой формы, длинных крыльев и короткого хвоста, а не за счет уменьшения веса. Приспособленность к тому или иному полету изменяет только скорость снижения, но не влияет на угол планирования. Тяжелый сапсан способен достигнуть такого же угла планирования, как и легкий, но при планировании он движется быстрее. Сглаженный контур минимизирует возмущения окружающего воздуха. Воздух гладко огибает крыло и, скручиваясь, срывается со свисающего края кончика крыла наподобие вихря (рисунок 1.15.2).
 Когда угол планирования становится круче, птица складывает крылья ближе к телу (рисунок 1.15.3) и летит быстрее. Это уменьшает рычажное давление на грудные мышцы, но слегка увеличивает тормозной эффект.
Когда угол планирования становится круче, птица складывает крылья ближе к телу (рисунок 1.15.3) и летит быстрее. Это уменьшает рычажное давление на грудные мышцы, но слегка увеличивает тормозной эффект.
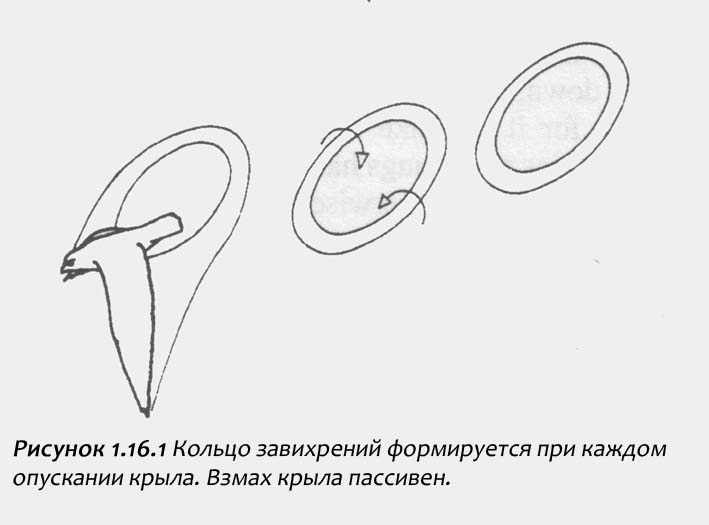 Забудем на некоторое время все воздушныe маневры типа поворотов, взлетов и приземлений. Активный полет, при котором ловчая птица продвигается вперед с помощью взмахов крыльев, имеет несколько узнаваемых типов. Первый - это размеренный полет, более или менее сравнимый с нашей ходьбой. Исследования Spedding, Pennycuick и Rayner показали, что когда сокол, например, пустельга или сапсан, летит очень медленно, со скоростью менее трех метров в секунду, движения крыльев вверх являются пассивными, не обеспечивающими подъема и не создающими вихрей. Результатом являются тороидообразные вихревые кольца, создаваемые толчками от движений крыльев вниз (рисунок 1.16. 1). При взмахе крыло сгибается очень близко к телу и поджимается, не создавая при этом следовых вихрей воздуха.
Забудем на некоторое время все воздушныe маневры типа поворотов, взлетов и приземлений. Активный полет, при котором ловчая птица продвигается вперед с помощью взмахов крыльев, имеет несколько узнаваемых типов. Первый - это размеренный полет, более или менее сравнимый с нашей ходьбой. Исследования Spedding, Pennycuick и Rayner показали, что когда сокол, например, пустельга или сапсан, летит очень медленно, со скоростью менее трех метров в секунду, движения крыльев вверх являются пассивными, не обеспечивающими подъема и не создающими вихрей. Результатом являются тороидообразные вихревые кольца, создаваемые толчками от движений крыльев вниз (рисунок 1.16. 1). При взмахе крыло сгибается очень близко к телу и поджимается, не создавая при этом следовых вихрей воздуха. Ястребы тоже способны к спринтерскому полету, но у них немного другая проблема. В то время как крупным соколам необходимо очень быстро преодолевать большие дистанции, ястребам нужно очень быстро летать на короткие дистанции. Они могут выиграть или проиграть схватку в считaнные секунды. Следовательно, у них проблемы не с наибольшей скоростью, а с ускорением. Ястребы начинают спринт со статического положения или во время медленного полета. Поэтому их спринт должен начинаться почти с нуля. В то время как кречет похож на велосипедиста, который выигрывает гонку на длинную дистанцию за счет максимальнай скорости, ястреб напоминает велосипедиста, который ожидает старта для гонки на 50 метров. Возможно, к финишу он даже не успеет набрать свою максимальную скорость. Следовательно, ему нужно полностью выложиться за короткий промежуток времени, поэтому он не может тратить половину этого времени на бесполезные поднятия крыла, которые не содействуют толчкам. Он решает эту проблему используя эластичные крылья с вырезками на первостепенных, которые не только аккумулируют энергию, но и уменьшают тормозной эффект при взмахе.
Ястребы тоже способны к спринтерскому полету, но у них немного другая проблема. В то время как крупным соколам необходимо очень быстро преодолевать большие дистанции, ястребам нужно очень быстро летать на короткие дистанции. Они могут выиграть или проиграть схватку в считaнные секунды. Следовательно, у них проблемы не с наибольшей скоростью, а с ускорением. Ястребы начинают спринт со статического положения или во время медленного полета. Поэтому их спринт должен начинаться почти с нуля. В то время как кречет похож на велосипедиста, который выигрывает гонку на длинную дистанцию за счет максимальнай скорости, ястреб напоминает велосипедиста, который ожидает старта для гонки на 50 метров. Возможно, к финишу он даже не успеет набрать свою максимальную скорость. Следовательно, ему нужно полностью выложиться за короткий промежуток времени, поэтому он не может тратить половину этого времени на бесполезные поднятия крыла, которые не содействуют толчкам. Он решает эту проблему используя эластичные крылья с вырезками на первостепенных, которые не только аккумулируют энергию, но и уменьшают тормозной эффект при взмахе. При условии активного полета в неподвижном воздухе и на одном уровне мы можем построить приближенныe кривые ускорения для различных хищников. Не имея возможности сравнить данныепо ловчим птицам, я скомпилировал эти кривые из нескольких источников, главным образом из работ Н. J. Slijper и Т. А. М. Jack, а также из собственных наблюдений. Хотя они, возможно, не очень точны для абсолютной скорости, они позволяют получить представление о различии между видами, хотя, конечно индивидуальные различия между птицами весьма значительны.
При условии активного полета в неподвижном воздухе и на одном уровне мы можем построить приближенныe кривые ускорения для различных хищников. Не имея возможности сравнить данныепо ловчим птицам, я скомпилировал эти кривые из нескольких источников, главным образом из работ Н. J. Slijper и Т. А. М. Jack, а также из собственных наблюдений. Хотя они, возможно, не очень точны для абсолютной скорости, они позволяют получить представление о различии между видами, хотя, конечно индивидуальные различия между птицами весьма значительны. далеко позади (например, куропатка). В таких случаях сарыч отстает и куда-нибудь садится, в данном случае на 45 метрах.
далеко позади (например, куропатка). В таких случаях сарыч отстает и куда-нибудь садится, в данном случае на 45 метрах. Лапы у хищных птиц выполняют семь функций: поддерживают вес тела; выполняют рулевые функции при некоторых маневрах; смягчают удар при приземлении; используются для хватания, удержания, а у некоторых видов и для умерщвления добычи; помогают регулировать температуру тела; используются для чистки недоступных для клюва частей тела и у некоторых видов используются при ухаживании. Для эффективного выполнения некоторых из этих функций лапы должны быть легкими для достижения максимальной проворности и иметь поверхность, которую легко очистить от крови, грязи и бактерий. Чтобы удовлетворять этим требованиям лапы эволюционировали до предела допустимых для живых тканей значений и допускают лишь очень небольшие отклонения. Поэтому неудивительно, что лапы очень подвержены заболеваниям.
Лапы у хищных птиц выполняют семь функций: поддерживают вес тела; выполняют рулевые функции при некоторых маневрах; смягчают удар при приземлении; используются для хватания, удержания, а у некоторых видов и для умерщвления добычи; помогают регулировать температуру тела; используются для чистки недоступных для клюва частей тела и у некоторых видов используются при ухаживании. Для эффективного выполнения некоторых из этих функций лапы должны быть легкими для достижения максимальной проворности и иметь поверхность, которую легко очистить от крови, грязи и бактерий. Чтобы удовлетворять этим требованиям лапы эволюционировали до предела допустимых для живых тканей значений и допускают лишь очень небольшие отклонения. Поэтому неудивительно, что лапы очень подвержены заболеваниям. а затем резко рванется, то обнаружится, что хватка ослабела. Естественно, храповик, как и все устройства, подвержен техническим проблемам; скопы часто тонут из-за того, что им не удается разжать хватку при нападении на слишком большую рыбу. Использование тонкой перчатки может побудить ловчую птицу, особенно легковозбудимую, вцепиться в руку. Таких птиц называют «прилипалами».
а затем резко рванется, то обнаружится, что хватка ослабела. Естественно, храповик, как и все устройства, подвержен техническим проблемам; скопы часто тонут из-за того, что им не удается разжать хватку при нападении на слишком большую рыбу. Использование тонкой перчатки может побудить ловчую птицу, особенно легковозбудимую, вцепиться в руку. Таких птиц называют «прилипалами». Некоторые виды соколов, такие как сапсан, для своего размера имеют большой вес, поэтому их лапы должны выдерживать большую нагрузку, чем у других. Крупныe соколы в неволе часто весьма малоподвижны, что увеличивает риск образования мозолей. Питание говядиной, похоже, увеличивает вероятность заболевания лап. Если дать птице разжиреть и кормить ее вечером, то всю ночь на лапы будут давить дополнительные 25% веса, как раз когда птица не активна и сидит на одной лапе.
Некоторые виды соколов, такие как сапсан, для своего размера имеют большой вес, поэтому их лапы должны выдерживать большую нагрузку, чем у других. Крупныe соколы в неволе часто весьма малоподвижны, что увеличивает риск образования мозолей. Питание говядиной, похоже, увеличивает вероятность заболевания лап. Если дать птице разжиреть и кормить ее вечером, то всю ночь на лапы будут давить дополнительные 25% веса, как раз когда птица не активна и сидит на одной лапе.