|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
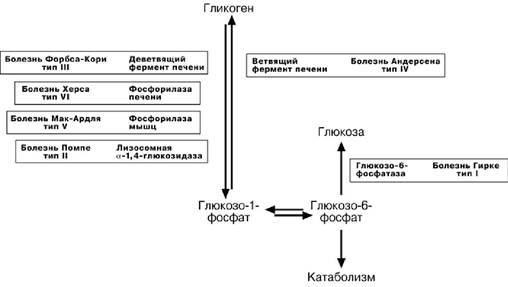
Печеночные гликогенозы
Самый частый гликогеноз I типа или болезнь фон Гирке обусловлен аутосом-
но-рецессивным дефектом глюкозо-6-фосфатазы. Из-за того, что этот фермент есть
только в печени и почках, преимущественно страдают эти органы, и болезнь носит еще одно название – гепаторенальный гликогеноз. Даже у новорожденных детей
наблюдаются гепатомегалия и нефромегалия, обусловленные накоплением гликоге-на не только в цитоплазме, но и в ядрах клеток. Кроме этого, активируется синтез липидов с возникновением стеатоза печени. Так как фермент необходим для де-фосфорилирования глюкозо-6-фосфата с последующим выходом глюкозы в кровь, у больных отмечается гипогликемия, и, как следствие, ацетонемия, метаболический ацидоз, ацетонурия.
Гликогеноз III типаилиболезнь Форбса-Кориилилимит-декстриноз–этоаутосомнорецессивный дефект амило-α-1-6-глюкозидазы, «деветвящего» фермен-та, гидролизующего α-1-6 -гликозидную связь. Болезнь имеет более доброкачествен-ное течение и частота ее составляет примерно 25% от всех гликогенозов. Для боль-ных характерна гепатомегалия, умеренная задержка физического развития, в подро-стковом возрасте возможна небольшая миопатия.
Еще два печеночных гликогеноза – гликогеноз IV типа (болезнь Андерсена),
связанный с дефектом ветвящего фермента и гликогеноз VI типа (болезнь Херса), связанный с дефицитом печеночной фосфорилазы гликогена встречаются довольно редко.
Мышечные гликогенозы
Для этой группы гликогенозов характерны изменения ферментов мышечной тка-
ни. Это приводит к нарушению энергообеспечения мышц при физической нагрузке, болям в мышцах, судорогам. Гликогеноз V типа (болезнь Мак-Ардля)–отсутствие мышечной фосфорила-
зы. При тяжелой мышечной нагрузке возникают судороги, миоглобинурия, хотя лег-кая работа не вызывает каких-либо проблем.
Смешанные гликогенозы
Эти заболевания касаются и печени, и мышц, и других органов.
Гликогеноз II типа (болезнь Помпе)–поражаютсявсегликогенсодержащие
клетки из-за отсутствия лизосомальной α-1-4-глюкозидазы. Происходит накопле-
ние гликогена в лизосомах и в цитоплазме. Заболевание составляет почти 10% всех гликогенозов и является наиболее злокачественным. Больные умирают в грудном возрасте из-за кардиомегалии и .тяжелой сердечной недостаточности.
Агликогенозы
Агликогенозы – состояния, связанные с отсутствием гликогена.
В качестве примера агликогеноза можно привести наследственный аутосомно-
рецессивный дефицит гликоген-синтазы. Симптомами является резкая гипогликемия натощак, особенно утром, появляется рвота, судороги, потеря сознания. В результа-те гипогликемии наблюдается задержка психомоторного развития, умственная от-сталость. Болезнь несмертельна при адекватном лечении (частое кормление), хотя и опасна.
ГЛИКОЛИЗ
Путь, по которому глюкоза окисляется до пировиноградной кислоты для получе-
ния энергии, называется гликолизом. В зависимости от дальнейшей судьбы пирува-
та различают аэробный и анаэробный гликолиз.
В аэробномпроцессе пировиноградная кислота превращается в ацетил-S-КоА и
далее сгорает в реакциях тканевого дыхания до СОB2B и НB2BО. В анаэробномпроцессе пировиноградная кислота восстанавливается до молоч-ной кислоты (лактата), поэтому в микробиологии анаэробный гликолиз называют молочнокислым брожением. Лактат является метаболическим тупиком и далее ни во
что не превращается, единственная возмож-ность утилизовать лактат – это окислить его обратно в пируват.
Практически все клетки организма способ-ны к анаэробному гликолизу. Для эритроцитов он является единственным источником энер-
гии. Клетки скелетной мускулатуры за счет бескислородного расщепления глюкозы спо- собны выполнять мощную, быструю, интенсив-
ную работу, как, например, бег на короткие дистанции, напряжение в силовых видах спор- та.
Анаэробный гликолиз локализуется в цито-золе и включает 2 этапа из 11 ферментатив-ных реакций.
Первый этап – подготовительный, здесь происходит
затрата энергии АТФ, активация глюкозы и образование
из нее триозофосфатов.
Первая реакциягликолиза,строго говоря,к гликоли-
зу не относится. Это гексокиназная реакция о которой
ранее уже говорилось ("Метаболизм гликогена"). Ее роль сводится к превращению глюкозы в реакционно способ- ное соединение за счет фосфорилирования 6-го, не
включенного в кольцо, атома углерода.
Для печени характерен изофермент гексокиназы –
глюкокиназа.Низкое сродство этого фермента к глюко-
зе обеспечивает ее захват печенью только после приема пищи, когда создается высокая концентрация глюкозы в крови. При обычных концентрациях глюкозы в крови пе-
чень ее не потребляет и та достается другим тканям. Вторая реакция–реакция изомеризации–необхо- дима для выведения еще одного атома углерода из
кольца для его последующего фосфорилирования. В ней образуется фруктозо-6-фосфат.
фосфата с образованием почти симметричной молекулы фруктозо-1,6-дифосфата. В четвертой реакции фруктозо1,6-дифосфат разре-
зается пополам с образование двух фосфорилирован-ных триоз-изомеров, альдозы глицеральдегида (ГАФ) и кетозы диоксиацетона (ДАФ).
Пятая реакцияподготовительного этапа–переходглицеральдегидфосфата и диоксиацетонфосфата друг в друга. Равновесие реакции сдвинуто в пользу диокси-
ацетонфосфата, его доля составляет 97%, доля глице-ральдегидфосфата – 3%. Эта реакция, при всей ее про- стоте, является вершителем судьбы глюкозы:
• при нехватке энергии в клетке и активации окис-
ления глюкозы диоксиацетонфосфат превраща-
ется в глицеральдегидфосфат, который далее окисляется на втором этапе гликолиза;
• при достаточном количестве АТФ, наоборот, глицеральдегидфосфат изомеризуется в диоксиацетонфосфат, и последний
отправляется на синтез жиров (см "Синтез триацилглицеридов").
Второй этап гликолиза – это освобождение
энергии, содержащейся в глицеральдегидфосфате,
и запасание ее в форме АТФ.
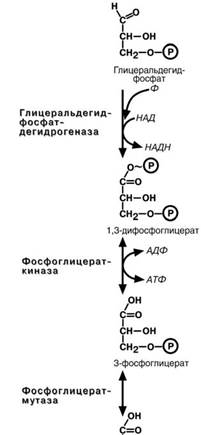
Шестая реакциягликолиза–окисление глице-
ральдегидфосфата и присоединение к нему фос-
форной кислоты приводит к образованию макроэр-гического соединения 1.3-дифосфоглицериновой кислоты.
В седьмой реакции энергия фосфоэфирной связи, заключенная в 1,3-дифосфоглицерате тра- тится на образование АТФ. Реакция получила до-
полнительное название –реакция субстратного фосфорилирования, что уточняет источник энергии для получения макроэргической связи в АТФ (суб-
страт) в отличие от окислительного фосфорилиро-вания (электрохимический градиент ионов водоро- да на мембране митохондрий).
Подобных реакций в клетке всего три – 1) ука-занная реакция, 2) пируваткиназная реакция, деся-тая реакция гликолиза (см ниже), 3) тиокиназная
реакция цикла трикарбоновых кислот.
Восьмая реакция–синтезированный в преды-
дущей реакции 3-фосфоглицерат изомеризуется в
2-фосфоглицерат.
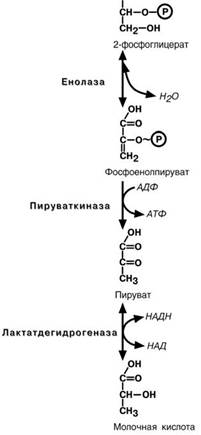
Девятая реакция–отрыв молекулы воды от
2-фосфоглицериновой кислоты приводит к образо-
ванию еще одной макроэргической фосфоэфирной связи. Еще одна реакция субстратного фосфорили-рования–десятая реакциягликолиза–заключа-
ется в переносе макроэргического фосфата с фос-
фоенолпирувата на АДФ.
Последняя реакция , одиннадцатая, образова-ние молочной кислоты из пирувата под действием лактатдегидрогеназы. Важно то, что эта реакция
осуществляется только в анаэробных условиях. Эта реакция необходима клетке, так как НАДН, об- разующийся в 6-й реакции, в отсутствие кислорода не может поступать и окисляться в митохондриях.
При наличии кислорода пировиноградная ки-
слота превращается в ацетил-S-КоА.
Поиск по сайту: |

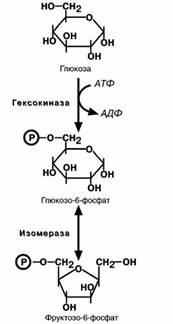
 Третья реакция–фосфорилирование фруктозо-6-
Третья реакция–фосфорилирование фруктозо-6-