|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Окислювальні процеси в клітинах⇐ ПредыдущаяСтр 27 из 27
Основним джерелом внутрішньоклітинної енергії е окислення субстратів киснем повітря. Але окислення в клітинах відбувається поступово, завдяки послідовному перенесенню
водню і електронів від субстрату до кисню. Окислення біосубстратів у живих організмах відбувається як в аеробних, так і в анаеробних умовах. Але до кінця (тобто з максимальним виділенням енергії з поживних речовин) окислювальні процеси відбуваються/тільки за участю атмосферного кисню. Процес, коли в реакціях біологічного окислення водень сполучається з киснем повітря, називається т к а -нинним диханням. Вивчення обміну білків, жирів і вуглеводів показало, що окислення цих сполук пов'язане з відщепленням водню , від субстратів, тобто їх дегідруванням. Окислення шляхом приєднання кисню до різних речовин відбувається, як правило, не за рахунок приєднання молекулярного атмосферного кисню, а за рахунок приєднання його із складу води. Поряд з водою другим кінцевим продуктом тканинного дихання є вуглекислий газ СОа, що утворюється під час процесів окислення органічних речовин при реакціях де-карбоксилювання. Основи сучасних уявлень про механізм тканинного дихання закладені в основному дослідженнями^ О. М. Баха та В. І. Палладіна, проведеними ще на початку XX ст.» а також дослідженнями В. Тунберга, О. Варбурга та ін. Всю систему тканинного дихання називають ще дихальним ланцюгом. Процес дихання відбувається у мітохондріях, які є енергетичними станціями клітини.
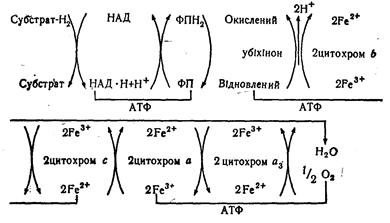
Дихальний ланцюг починається з ферментів дегідроге--наз, що каталізують дегідрування субстратів (див. с. 314). Вони відрізняються специфічністю за відношенням до субстратів і завжди мають у своєму складі коферменти. Існує дві групи дегідрогеназ, які розрізняються за вмістом коферментів: одні мають у своєму складі нікотинамідаденін-динуклеотид (НАД), інші — флавінмононуклеотиди (ФМН) або флавінаденіндинуклеотид (ФАД). Тобто існують НАД- або ФАД-залежні дегідрогенази. Ці два типи дегідрогеназ мають різні окислювально-відновні потенціали залежно від природи субстратів, що ними використовуються. НАД-Залежні дегідрогенази е первинними акцепторами атомів водню, тобто вони здатні віднімати протони й електрони безпосередньо від субстратів. Флавінові коферменти зв'язані з вторинними дегідрогеназами, що відщеплюють тільки водень від відновленої форми НАД, тобто найважливішою дегідрогеназою є НАД • Н-дегідрогеназа. Друга важлива дегідрогеназа — сукцинат-Дегідрогеназа, що безпосередньо відщеплює водень від субстрату сукцинату в лимоннокислому циклі. Деякі ФАД-залежні дегідрогенази здатні передавати водень безпосередньо кисню, тобто є оксидазами. Проміжною сполукою в клітинному окисленні в убіхі-нон, або кофермент0, що був виділений із ліпідів міто-хондрій. У дихальний ланцюг входять також цитохроми — хромопротеїди, що здатні переносити електрони завдяки наявності в їх складі заліза, яке може переходити із тривалентного у двовалентне. Нині вивчено вже кілька десятків цитохромів. Чим нижчий окислювально-відновний потенціал переносника електронів або водню, тим кращим він є відновником і тим ближче він знаходиться до субстрату. Сполука може віддавати свій водень або свої електрони іншій сполуці, що має вищий потенціал. Отже, порядок розташування ферментів і коферментів у дихальному ланцюгу такий:
Наведені значення окислювально-відновного потенціалу орієнтовні тому, що для НАД і флавіннуклеотидів вони«мі-нюються залежно від природи апоферменту.
^ Окислювальний ланцюг може бути представленийтакоюсхемою:
Безпосереднє окислення субстрату відбувається аа рахунок ферменту, у якого кофермент (переносник водню) має окислювально-відновний потенціал, що близький до потенціалу субстрату. Синтез макроергічного зв'язку АТФ або АДФ потребує 30,6 кДж/моль енергії, що відповідає реакції, в якій беруть участь два електрони (2Н+ або 2е~) при зміні окислювально-відновного потенціалу приблизно на 0,16 В. Найбільші зміни потенціалу відбуваються при переході від НАД до ФАД (або ФМН), від убіхінону до цитохрому с і між ци-тохромом а і киснем. Кожний з цих переходів відповідає утворенню однієї молекули АТФ з АДФ і фосфату. Отже, окислення НАД . Н + Н+ дає можливість утворитися трьом, а окислення ФАД • Н, — двом макроергіч-ним зв'язкам. Окислення іонів Ре^"*" у цитохромах забезпечує синтез одного такого зв'язку. Тобто окислення субстрату ферментом, який має в своєму складі НАД+, у присутності кисню веде до утворення трьох молекул АТФ. У результаті окислення субстрату ферментом, що має в своєму складі ФАД, утворюється дві молекули АТФ, а в результаті окислення субстрату тільки цитохромною системою однієї молекули АТФ. Кількісний зв'язок між синтезом АТФ і використанням аисню можна охарактеризувати відношенням Р/0, яке вказує на кількість атомів фосфору, що перейшла з енергетичного стану в молекулу АТФ ^іа кожну 1/, молекули кио" ню. Це відношення дорівнює трьом для НАД • Н + Н+ і двом — для ФАД • Нд. Отже, клітинне дихання супроводжується фосфорилюванням АДФ. Це явище називається окислювальним фосфорилюванням. В організмі має місце спряженість між окисленням і фосфорилюванням. Макроергічні зв'язки АТФ можуть утворюватися за рахунок субстратного фосфор илювання, що має місце в двох випадках в гліколітичному циклі (при окисленні глі-церальдегід-3-фосфату до 3-фосфогліцеринової кислоти і при перетворенні 2-фосфогліцеринової кислоти у піровиноградну). Інтенсивність дихання регулюється відношенням АТФ/АДФ. Чим менше це відношення, тим інтенсивніше відбувається дихання, забезпечуючи утворення АТФ. Підвищення рівня АТФ знижує дихальні процеси. Агенти, що роз'єднують утворення АТФ (динітрофенол, тироксин), активують дихання. За останні роки одержано ряд нових даних, що розкривають деякі особливості структури і функції дихального ланцюга. Так, детальніше висвітлена роль мітохондрій в процесах дихання і трансформації енергії. Відповідно за хеміосмотичною теорією П. Мітчела мембрани здатні трансформувати хімічну енергію окислення в енергію мембранного потенціалу (тобто електричну), а синтез АТФ відбувається за рахунок розряджування мембран за участю ферменту протонної аденозинтрифосфатази. Мембрана з її електричним потенціалом і є тим джерелом, яке не враховували раніше при дослідженнях ферменту, що відповідає за біосинтез АТФ (окислювальне фосфорилювання). Енергія мембранного потенціалу використовується[також для транспорту іонів проти градієнта концентрації, а також для утворення теплоти. Більша частина енергії, резервованої в АТФ, використовується для забезпечення потреб організму. Діяльність скелетних м'язів, м'язів серця, нервової системи, рух шлунку, перистальтика кишок, всмоктування речовин з кишок, робота нирок, легенів та інші життєві функції забезпечуються енергією макроергічних зв'язків, переважно АТФ. Для біосинтезу різних субстратів використовується в середньому близько 10—15 ''% енергії, тобто основна маса енергії використовується на утворення теплоти тіла і на забезпечення його функцій і значно менша кількість — на відтворення тканин.
Пути образования АТФ.
В настоящее время в основных чертал известны все пути, ведущие к образованию АТФ, хотя детали самого процесса аккумулирования энергии в молекуле АТФ не ясны. В любом случае, независимо от того, каким путем это происходит, процесс образования АТФ называется фосфорилированием. Основным условием фосфорилирования, т. е. образования АТФ, является наличие окислительно-восстановительных процессов. Механизмы этих процессов существенно различаются в зависимости от использования энергии солнечного света (у автотрофов) или энергии окисления органических веществ (у гетеротрофов). Однако общим для обоих случаев является то, что в основе окислительно-восстановительных процессов лежит перенос электронов и водорода. При отсутствии света автотрофные организмы существуют за счет тех же энергетических и конструктивных процессов, что и гетеротрофные. , Автотрофный обмен — источник жизни на Земле. Используя энергию солнечного света, автотрофы синтезируют органические вещества из оксида углерода (IV). Процесс образования органических веществ из оксида углерода (IV) под влиянием солнечного света называется фотосинтезом. Гетеротрофы существуют за счет веществ, накопленных в процессе фотосинтеза. В результате фотосинтеза образуется и кислород, необходимый для жизнедеятельности гетеротрофов. Таким образом, процесс образования веществ при фотосинтезе—это своеобразный способ запасания (консервирования) солнечной энергии, которая используется затем и ге-теротрофами, и автотрофами. Солнечная энергия, запасенная в органических веществах, используется во всех видах человеческой дея тельности. Сжигание топлива—угля, нефти, газа, торфа, древесн ны — является основным источником получения всех видов энергии. Химизм процесса фотосинтеза сложен и не до конца изучен. Достаточно хорошо известен химизм превращения углекислого газа в органические вещества. Менее изучена биохимия процесса преобразования солнечной энергии в 'энергию макроэргических связей АТФ. По существующему представлению, процесс фотосинтеза у растений и фотосинтезирующих микроорганизмов протекает в две фазы: световую и темновую. В световой фазе под влиянием света образуются АТФ и восстановленные дегидрогена-зы. В темновой фазе (при отсутствии света) из углекислого газа с помощью АТФ и восстановленных дегидроненаз синтезируются органические вещества. Принципиальный механизм образования АТФ у растений и фотосинтезирующих микроорганизмов один и тот же. Движущей си-
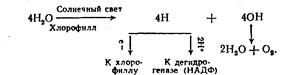
лой фотофосфорилирования в обоих случаях является изменение окислительно-восстановительного потенциала, вызванное -поглощением света фоточувствительными системами (фотосенсибили-заторами) и сопровождающееся перемещением электронов и водорода. Этот принцип лежит и в основе окислительного фосфори-лирования, т. е. энергетического обмена у гетеротрофов, только в этом случае разность потенциалов и перемещение электронов и водорода создают дегидрогеназы, взаимодействуя с окисляемым субстр^гом^Яринцин фотофосфорилирования заключается в следую-/Щем. Система фотосенсибилизаторов поглощает световые лучи и ^ изменяет свой окислительно-восстановительный потенциал. Различные по химической природе фотосенсибнлизаторы поглощают ? лучи с различной длиной волны. Вследствие этого между система-» ми появляются разность потенциалов и поток электронов. Это ., сопровождается выделением энергии, за счет которой АДФ превращается в АТФ, т. е. идет процесс фотофосфорилиро-; в а н и я. Фоточувствительные системы сложны. Все они содержат 1 пигменты. Главные из них — хлорофилл у растений и бактерио-хлорофилл у микроорганизмов. Кроме хлорофилла в процессе фотосинтеза участвуют каротиноиды, хиноны (пластохинон у рас-• тений и убихинон у микроорганизмов), ферредоксин, цитохромы | /I, Ь, с, /. Способностью поглощать солнечные лучи обладают лишь ! хлорофилл и некоторые другие пигменты, остальные вещества яв-' ляются просто передатчиками электронов на восстановленные дегидрогеназы. Первым в цепи передачи электронов всегда стоит хлорофилл. Конечный акцептор электронов — дегидрогеназа. Таким образом, процесс передачи электронов приводит к образованию не только АТФ, но и восстановленной дегидрогеназы. Каждый посредник, отдавая электроны, восполняет их потерю за счет электронов предыдущего передатчика, т. е. переходит в первоначальное (восстановленное) состояние. В противном случае, т. е. при безвозвратной отдаче электронов, расходовались бы пигменты. Хлорофилл же, стоящий первым в цепи передачи, восстанавливает утраченные электроны за счет электронов водорода (у растений для этого используется водород воды, у микроорганизмов — водород сероводорода или других соединений). Сам же водород при этом передается дегидрогеназе. Процесс мобилизации водорода и электронов у растений осуществляется в результате разложения воды под влиянием солнечного света (фотолиза), происходящего с участием хлорофилла:
В результате фотолиза воды образуется свободный кислород, используемый гетеротрофамй и самими растениями для окисления органических веществ и швлечеиия из них энергии. Таким образом, растения, запасая солнечную энергию в виде органических веществ, запасают при этом и кислород, необходимый для извлечения этой энергии. Процесс мобилизации водорода и электронов у фотосинтези-рующих серобактерий можно представить схемой
Вместо свободного О; у микроорганизмов в данном случае образуется молекулярная 5, которая откладывается в их клетках. Растения восполняют утраченные хлорофиллом электроны и другим способом, при котором, пройдя цепь переносчиков, электроны вновь возвращаются к хлорофиллу. Этот способ тоже приводит к образованию АТФ и называется циклическим фос-форилированием. Однако восстановленные дегидрогеназы при этом не образуются. Таким образом, в основе всех способов превращения световой энергии в биологическую лежит один и тот же принцип, различие состоит лишь в источниках и путях передачи электронов и водорода. Ниже показаны основные системы, участвующие в процессе переноса электронов и водорода при фотофосфор ил ирова и и и у растений (а) и фотосинтезирующих микроорганизмов (пурпурных серобактерий) (б), использующих энергию света:
В настоящее время еще многое не ясно о химизме фотосинтеза. Роль ряда веществ, участвующих в этом процессе, не установлена. Не ясен вопрос интенсивности фосфорилирования и количества образующихся молекул АТФ. Известно, что у растений АТФ образуется в момент передачи электронов от водорода воды к хлорофиллу, затем — от цитохрома Ь к. цитохрому /. У микрооргапиз-
Умов эти стадии пока не определены, хотя нет сомнения в том, что ^передача электронов приводит к образованию АТФ. Установлено, что передача водорода от восстановленной дегид-рогеназы к фосфоглицернновой кислоте также сопровождается образованием АТФ, однако это уже не относится к световой фазе. Извлечение энергии из органических веществ при гетеротрофном обмене основано также на принципе переноса электронов и водорода. Источником электронов и водорода при этом является органическое вещество (окисляемый субстрат), роль переносчиков выполняют дегидрогеназы и цитохромы. Акцептор водорода и электронов — кислород воздуха, а при отсутствии воздуха акцептором может служить кислород каких-либо минеральных веществ. На основании многочисленных работ сформулирована теория окисления веществ в живом организме, называемая теорией биологического окисления (теорией дыхания). Согласно этой теории, окисление вещества начинается с отнятия водорода пиридиновыми дегидрогеназами и передачи его флавиновым дегидрогеназам. После этого система цитохромов передает электроны этого водорода кислороду. В результате присоединения электронов кислород переходит в ионную форму. Водород, отдавший электроны, также ионизируется и переходит в раствор. Заряженные частицы водорода н кислорода соединяются между собой, образуя воду. Ниже представлена упрощенная схема переноса На и электронов при окислительном фосфорилировании (дыхательная цепь):
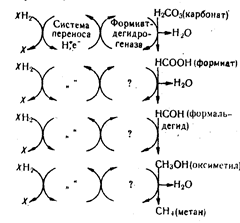
На схеме показаны только четыре цитохрома: Ь, с, а и из. участие которых наиболее изучено (ХНг—окисляемый субстрат). В дыхательной цепи участвуют и другие вещества. Большинство из них играют вспомогательную роль и не являются непосредственными переносчиками протонов и электронов. Есть также и вещества небелковой природы, например, убихинон, стоящий в цепи между флавиновой дегидрогеназой и системой цитохромов. Биологический смысл переноса водорода и электронов заключается в использовании энергии образования воды. Реакция соединения водорода и кислорода—наиболее эффективная экзотермическая реакция между химическими элементами. Природа использовала эту реакцию в качестве источника энергии для биохимических процессов. Вне организма эта реакция происходит мгновенно. со взрывом, а в живом организме — постепенно. Водород передается кислороду через цепь передатчиков, и энергия выделяется не сразу, а по частям, что дает возможность использовать ее для биохимических процессов. Каждый этап перемещения водо рода и электронов от фермента к ферменту сопровождается выделением энергии, и на определенных этапах выделяющейся энергии достаточно для осуществления процесса фофорилирования и образования молекулы АТФ. Во всей цепи установлено три таких этапа. Процесс образования АТФ при биологическом окислении называется окислительным фосфорилированием. Этот путь образования АТФ является универсальным и используется всеми организмами в процессе их жизнедеятельности в присутствии кислорода. Многие микроорганизмы, в том числе и промышленные, способны существовать при отсутствии кислорода. В этом случае они используют иные пути образования АТФ. Один из них близок к универсальному пути биологического окисления. Отличие состоит в том, что при этом микроорганизмы используют в качестве конечного акцептора водорода кислород окисленных минеральных веществ, например сульфатов, нитратов. В результате, как и в общем случае, образуется вода, т. е. организм использует энергию образования воды. Такие процессы называют редукцией. Процесс использования кислорода может быть частичным или полным. При этом кислород одной молекулы вещества неоднократно акцептирует водород окисляемого субстрата. Схема процесса переноса водорода и электронов на кислород нитратов (а) и сульфатов (б) приведена ниже:
Детали процесса переноса водорода и электронов к кислороду указанных акцепторов пока еще не ясны и не известно количество образующихся при этом молекул АТФ. Предполагают, что нитрат-редуцирующие бактерии используют те же ферменты переноса водорода, которые осуществляют этот процесс в присутствии кислорода воздуха, только вместо цитохромоксидазы конечным переносчиком является нитратредуктаза. Цитохромы же у микроорганизмов, существующих в анаэробных условиях, не обнаружены. Более подробно изучен процесс восстановления сульфатов. Известно, что в нем участвует фермент АТФ—сульфатаденилил-трансфераза (АТФ-сульфурилаза). Под действием этого фермента образуется аденозин-З^фосфосульфат, который под влиянием специальной редуктазы превращается в сульфат и АМФ. Далее идет восстановление до сульфида, но детали этого процесса не ясны. Учитывая, что микробиологические процессы, для которых характерна редукция нитратов и сульфатов/всегда сопровождаются образованием метана, можно предположить, что параллельно с этими процессами идет редукция карбонатов, кислород которых также является акцептором водорода окисляемых веществ в анаэробных условиях. Карбонаты восстанавливаются аналогично нитратам и сульфатам. Конечным продуктом восстановления является метан. Схема процесса переноса водорода и электронов на кислород карбонатов приведена ниже: В литературе этот процесс рассматривается как восстановление СОа специальными метанобразую-щими бактериями. Причем для восстановления эти бактерии используют водород, отщепляемый от органических веществ другими микроорганизмами. Однако более вероятно, что мобилизация водорода окисляемых ве-I) ществ и передача его к кислороду карбонатов — это, как и в случае редукции нитратов и сульфатов, единый окислительно-восстано-
Вг.тельный процесс, происходящий в одном и том же организме. Первый продукт восстановления—формиат—образуется, как известно, с участием формиатдегидрогеназы. Аналогичную роль выполняют нитрат- и сульфатдегидрогеназы. Вполне вероятно, что кислород нитратов, сульфатов н карбонатов в анаэробных условиях может использовать один и тот же микроорганизм. Второй путь образования АТФ — субстратное фосфорилирова-ние. Оно также используется организмами при отсутствии кисло рода воздуха. Осуществляется не вследствие окислительно-восстановительного процесса переноса водорода и электронов, а прямым переносом остатка фосфорной кислоты от какого-либо фосфори-лированного вещества (субстрата) на АДФ с помощью ферментов киназ. Этот путь используют в качестве единственного источника АТФ различные виды промышленных микроорганизмов—дрожжей и бактерий в процессе брожения (в присутствии кислорода они переходят на аэробное окисление органических веществ). В аэробных условиях субстратное фосфорилирование является дополнительным источником АТФ у всех организмов, потребляющих углеводы. При разложении других веществ субс1 ратное фосфорилирование не происходит. Источниками фосфата в этом случае являются фосфоглицериновая и фосфоэнолпиропшюгрпдпая кислоты.
Поиск по сайту: |