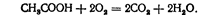|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Цикл трикарбонових кислот
Цикл трикарбонових кислот було відкрито в 1937 р. Р. Кребсом, і тому він одержав назву циклу Кребса. Оскільки першою сполукою, що утворюється в ньому, є ^іимонна кислота, то він ще називається циклом лимон-йої кислоти, або цитр^тним циклом. Цикл починається з реакції конденсації ацетил-КоА в оксалоацетатом (щавлевооцтова кислота — ЩОК), 'внаслідок чого утворюється цитрат (лимонна кислота). Ця реакція каталізується ферментом цитратсинтетазою (див. с. 249). Внаслідок цієї реакції ацетильна група ацетил-КоА приєднується до вуглецю карбонільної групи оксало-ацетату з наступним гідролізом тіоефірного зв'язку і утворенням вільного КоА —8Н. Цитратсинтетаза належить до регуляторних ферментів, її активність знижується при значних кількостях АТФ або НАД. Далі лимонна кислота (цитрат) під дією ферменту аконі-татгідратази (аконітази) дегідрується і перетворюється на ((мс-аконітову кислоту (({иоаконітат), що після приєднання молекули води перетворюється в ізолимонну (ізоцитрат). Фермент аконітатгідратаза фактично забезпечує досягнення рівноваги в такій системі: лимонна кислота, цис-аконітова кислота, ізолимонна кислота. При рН == 7,5 і 25 °С в системі знаходиться 91 % лимонної, 6 % іго-лимснної і 3 % цис-аконітової кислот. Фермент фактично здатний каталізувати приєднання води за місцем подвійних зв'язків цис-аконітату одночасно в двох напрям- ' іах — з утворенням або цитрату, або ізоцитрату. Наступні перетворення відбуваються через ізолимонну кисДрту, а її запаси поповнюються за рахунок ізомеризації лимонної кислоти через ((Ус-аконітову. Ізо цитрат окислюється до а-кетоглутарату відщепленням двох атомів водню і декарбоксилюванням. Каталізує цю реакцію фермент ізоцитратдегідрогеназа, коферментом якого є НАД4'. На цьому етапі починається окислення
Цикл трикарбонових кислот; І — цитратсинтаза; і — аконітатгідратаза! З — Іаоцитратдегідрогеназа? 4 — а-кетоглутаратдегідрогеназннй комплекс; 5 •«• сукцинілтіокіназа; 6 « вукцинатдегідрогеваза) 7 — фумаратгідратааа; Й — малатдегідрогеназа залишку оцтової кислоти і перетворення трикарбонових кислот у дикарбонові. Ізоцитратдегідрогеназа є алостеричним (регуляторним] ферментом, активність якого дуже знижується в присутності значних кількостей АТФ і НАД • Н. При підвищенні концентрації АДФ у клітині збільшується швидкість окислення ізоцитрату ферментом, а разом з тим і швидкість всього циклу Кребса тому, що саме ця реакція лімітує швидкість процесу в цілому. Підвищення швидкості реакцій у циклі Кребса викликає прискорений
перенос електронів і, як наслідок, підсилення окислювального фосфорилювання, тобто перетворення АДФ у АТФ. З підвищенням концентрації АТФ кількість АФД зменшується, що веде до «виключення» ізоцитратдегідро-генази. Накопичення НАД • Н у мітохондріях також знижує окислення ізоцитрату. Так відбувається автоматична регуляція активності ферментів у циклі Кребса. На наступному етапі відбувається окислення а-ке-тоглутарату до сукцинату (янтарної кислоти). Цей процес. проходить у дві стадії і є необоротним. Спочатку . відбувається окислювальне декарбоксилювання а-кето-глутарату, внаслідок чого утворюється сукциніл-5 — КоА. Ця реакція подібна до реакції окислення пірувату з утворенням ацетил-КоА. Каталізує цю реакцію глутамат-дегідрогеназний комплекс, який дуже схожий на піруват-дегідрогеназний комплекс як за будовою, так і за механізмом дії. Кінцевий продукт цієї реакції сукциніл-КоА є високоенергетичним тіоефіром, в утворенні складноефірного зв'язку якого бере участь одна з карбоксильних груп янтар-взї кислоти. На наступному етапі сукциніл-КоА взаємодіє з гуано-зиндифосфатом (ГДФ) і неорганічним фосфатом. При цьому енергія, що знаходилася у макроергічному зв'язку сукциніл-КоА, переходить у фосфатний зв'язок ГТФ. Кофермент КоА —8Н вивільнюється в реакції. - Ця реакція каталізується ферментом сукцинілтіокіназою (сукциніл-КоА-синтетазою), що каталізує утворення високоенергетичного фосфатного зв'язку ГТФ за рахунок високоенергетичного тіоефірного зв'язку сукциніл-5 — КоА. ГТФ, що утворилася в результаті цієї реакції, віддає потім свою кінцеву фосфатну групу АДФ, внаслідок чого утворюється АТФ:
Синтез АТФ після деацилювання сукциніл-5 —КоА називається ще фосфорилюванн.ім на рівні субстрату або субстратним фосфорилюванням, щоб відрізнити його від реакцій фосфорилювання, що проходять за участю дихального ланцюга. Отже, при перетворенні ізолимонної (трикарбонової) кислоти у янтарну (дикарбонову) відщеплюється дві молекули СОз, тобто окислюється два атоми вуглецю, що входили до складу ацетильної групи. Сукцинат (янтарна кисло та) окислюється до фумарату (фумарової кислоти) за участю ферменту сукцинатдегідрогенази, коферментом якого є флавінаденіндинуклеотид (ФАД). Відновлюючись, він переходить у ФАД • На, який далі окислюється у дихальному ланцюгу за участю цитохромів. Фумарат під час гідратації за участю ферменту фу-маратгідратази утворює а-малат (яблучну кислоту). В останній реакції циклу трикарбонових кислот ма-Латдегідрогеназа при наявності НАД4" окислює малат, відщеплюючи водень, в оксалоацетат (щавлевооцтову кислоту), з якого почався цикл трикарбонових кислот. Отже, в циклі трикарбонових кислот відщеплення водню відбувається у чотирьох випадках: при окисленні ізолимонної кислоти в ос-кетоглутарову; окислювальному декарбоксилюванні а-кетоглутарової кислоти з утворенням сукциніл-КоА; окисленні фумарової кислота до яблучної та яблучної до щавлевооцтової. Всього відщеплюється чотири молекули водню. У циклі Кребса було приєднано дві молекули води (4 атоми водню) — при розриві комплексу лимонної кислоти з КоА і при перетворенні фумарової кислоти в яблучну. Тому за рахунок кспалювання» ацетильної групи вивільнюється дві молекули водню, тобто весь водень, що був у молекулі. Декарбоксилювання проходить у двох випадках — при перетворенні ізолимонної кислоти в а-кетоглутарову і а-кетоглутарової кислоти в янтарну. В циклі трикарбонових кислот ацетильний залишок окислюється з утворенням двох молекул Н^О і двох молекул СОд. Для його окислення використовується дві молекули кисню. Загальне рівняння має такий вигляд:
Отже, на одну молекулу СОд витрачається одна молекула кисню, тобто СОа/Оз =° 1. Співвідношення кількості СОд, що виділився за певний час, до кількості кисню, що поглинутий за той самий час, називається дихальним коефіцієнтом. При обміні вуглеводів він дорівнює одиниці. Частина кисню, що необхідна для утворення двох молекул СОд, забезпечується за рахунок двох реакцій гідратації. У процесі дихання основним є відщеплення атомів водню (дегідрогенізація); декарбоксилювання є підпорядкованою ланкою і обумовлюється інтенсивністю
дегідрогенізації. У циклі Кребса утворюється чотири пари атомів водню: три пари використовуються для відновлення НАД'1', а одна — для відновлення ФАД сукцинат-дегідрогенази. Усі чотири пари атомів водню перетворюються в іони Н4" і відповідна кількість електронів переноситься по дихальному ланцюгу на кисень. Отже, існує зв'язок між ферментами циклу Кребса< окислювальними ферментами і процесами окислювального фос-форилювання. Ферменти циклу Кребса локалізовані в матриксі мітохондрій поряд з ферментами дихального ланцюга, що знаходяться у внутрішній мембрані. В аеробній фазі окислюється від 1/5 або 1/6 до 1/3 частини молочної кислоти залежно від потреб організму. З решти молочної кислоти за рахунок енергії дихання знову синтезується глікоген, який резервується в клітині і використовується за потребою. Значення циклу трикарбонових кислот для організму полягає не тільки в забезпеченні організму енергією, айв утворенні проміжних речовин, що використовуються під час біосинтезу: піровиноградна кислота, ацетил-КоА, а-кетоглутарова кислота тощо. Фотосинтез Біологічне значення фотосинтезу дуже важливе, оскільки за допомогою молекулярних механізмів цього процесу сонячна енергія вловлюється фотосинтезу'ючими клітинами і перетворюється в хімічну. Сонячна енергія є джерелом усіх видів енергії, які використовуються в біологічних системах. Здатність до фотосинтезу мають різні організми — як прокаріоти, так і еукарїоти. До фотосинтезуючих еукаріотів належать вищі зелені рослини, а також нижчі форми — багатоклітинні зелені, бурі і червоні водорості, одноклітинні організми — евгленові, дінофлагелляти і діатомові водорості. До фотосинтезуючих прокаріотів належать синьо-зелені водорості, а також зелені і пурпурові бактерії. Фотосинтез на земній поверхні мабуть більше, ніж наполовину обумовлюється саме одноклітинними організмами, особливо водоростями, такими як діатомові і дінофлагелляти. Фотосинтезом називається синтез, що проходить з використанням світлової енергії. В живій природі він локалізований у спеціальних органелах рослинних клітин — хлоропластах, що подібні до мітохондрій тваринних клітин як у відношенні мікроструктури, так і у відношенні молекулярної організації. Обидва типи органел мають специфічні форми ДНК і здатні до самовідтворення, яке відбувається певною мірою незалежно від хромосом ядра. Хлоропласти мають у своєму складі кілька забарвлених пігментів, хлорофілів, що зв'язані з білками і ліпідами. Своєю тетрапірольною конфігурацією вони подібні до гему гемоглобіну, але замість заліза в цій сполуці є магній-Фотосинтез — процес; оборотний диханню, бо в ньому відбувається фіксація СО^ і виділення кисню. У всіх фото синтезуючих організмів, крім бактерій, вода є донором електронів або атомів водню для відновлення СОд:
Газовий обмін супроводжується синтезом вуглеводів. Енергія поглинутого світла фотосинтезуючими системами викликає потік електронів від донорів до акцепторів електронів. Напрямок загального потоку електронів під час фотосинтезу завжди протилежний напрямку нормального градієнта стандартного відновлювального потенціалу і відповідає їх руху в напрямку'більш електронегативних систем. Енергію, необхідну для переміщення електронів, поставляють кванти світла. За участю світла (світлова стадія) відбувається відновлення НАДФ+ воднем з молекул води і фосфор илювання АДФ. У темновій стадії відновлена форма НАДФ • Н і АТФ використовуються для відновлення СС>2 до гексози. Фотосинтезуючі клітини мають три типи пігментів, що вловлюють кванти світла: хлорофіли, каротиноїди і фікобі-ліни. Більшість фотосинтезуючих клітин, що виділяють кисень, мають два різних хлорофіли: один (обов'язково) хлорофіл а, а другий — хлорофіл Ь (у зелених рослин), хлорофіл с (у бурих водоростей), хлорофіл гі (у червоних водоростей). Усі хлорофіли дуже добре поглинають видиме світло, що обумовлюється наявністю великої кількості у них спряжених подвійних зв'язків у молекулах. Енергія поглинання фотонів при цьому значною мірою делокалізує-ться, розподіляючись по всій електронній структурі молекули хлорофілу. Під час фотосинтезу енергія світла, кількість якої залежить від довжини хвилі світла, поглинається пігментами, які переходять при цьому у збуджений стан, що характеризується більшою енергією. Енергія збудження вловлюється спеціальною молекулою пігменту, що віддає свій електрон, який використовується для відновлення НАДФ+. У клітинах, що виділяють кисень, є дві фотосистеми. Фо-тосистема І має хлорофіл а і р-каротин, а також одну молекулу пігменту Р;а„, ЩО уловлює енергію поглинання світла: ця молекула активується переважно світлом з більшою довжиною хвиль. Пігмент Руоо знаходиться в дуже малих кількостях і має максимум поглинання при довжині хвилі 700 нм. Вважають, що пігмент Руоо — це спеціалізована форма хлорофілу:
Фотосистема II має інший набір пігментів. Вона активується світлом з меншою довжиною хвилі і відповідає за виділення кисню. Фотосистеми І і II об'єднані в агрегат. Збудження фотосистеми І підвищує потенціал електрона до дуже високого значення, достатнього для відновлення НАДФ"1", але попередньо електрон проходить через ланцюг переносників, які «включають» залізовмісний білок фередо-ксин і фермент фередоксин-НАДФ+'оксидоредуктазу. Електрони поступають з молекул води, які віддають водень, механізм цього явища ще невідомий. Світлова енергія одночасно з відновленням НАДФ+ воднем води забезпечує синтез АТФ. Процес фосфор илю-вання АДФ, спряжений з одним або більше ланцюгів переносу електронів, називається фотофосфорилю-в а н н я м. Фотофосфорилювання мабуть має багато спільного з окислювальним фосфорилюванням. Для пояснення процесу фотофосфорилювання запропоновані механізми, що базуються на хімічному, хімікоосмотичному і конформа-ційному спряженнях. Для виділення однієї молекули кисню і відновлення однієї молекули СОд необхідно вісім квантів світла з довжиною хвилі 700 нм. При цьому утворюються дві молекули НАДФ • Н + Н+ і три молекули АТФ;
Кінцевим продуктом фотосинтезу є залишки глюкози. Синтез глюкози відбувається у темновій стадії, де енергія накопичена у вигляді фосфатних зв'язків АТФ і НАДФ • Н, що використовуються для відновлення СО,. При біосинтезі глюкози (або гексози) спочатку проходить карбоксилювання і гідролітичне розщеплення рибулозо-1,5-дифосфату з утворенням двох молекул 3-фосфогліцери-нової кислоти. Каталізує цю реакцію фермент дифосфори-булозокарбрксилаза або рибулозодифосфаткарбоксидисму- тазаі
Відновлення 3-фосфогліцеринової кислоти відбувається за участю ферменту гліцеральдегід-3-фосфатдегідрогенази, що має в своєму складі НАДФ • Н. Гліцеральдегід-3-фосфат під дією ферменту тріозофосфатізомерази частково перетворюється в діоксіацетонфосфат і за допомогою альдолази використовується під час гліколізу для синтезу глюкози. Гліцеральдегід-3-фосфат через пентозофосфатний (фосфо-глюконатний) цикл, коли реакції проходять у зворотному напрямку, утворює рибулозо-5-фосфат, який, перетворюючись у рибулозо-1,5-дифосфат, повертається в цикл фото синтезу. Послідовність реакцій утворення вуглеводів у листках рослин при фотосинтезі можна зобразити такою схемою:
Утворення глюкози з СОа при фотосинтезі через цикл Кальвіна (скорочення): ЗФГ — З-фосфогліцеринова кислота; ГЗФ — гліцеральдегід-3-фосфат. ДОАФ - діоксіацетонфосфат; Ф6Ф - фруктозо-б фосфат; ФДФ-фрукто зо-І 6-дифосфат; Г6Ф - глюкозо-6-фосфат; Е4Ф - еритрозо-4-фосф^т; Кс5Ф -ксилулозо-Б-фосфат; СДФ - седогептулозо-1.7-дифосфат, С7Ф -«едогептулозо-7-фосфат; Р5Ф - рибозо-6-фосфат; Ру5Ф - рибу лозо 5 фосфат; РуДФ — рибу лозо-1,5-дифоеф<ї Сумарна реакціяі
Цей цикл називається циклом Кальвіна і включає перетворення вуглеводів як за участю ферментів гліколізу, так і частково за участю ферментів фосфоглюконатного циклу (див. с. 266) Деякі реакції мають самостійне значення. З глюкози в листках рослин синтезуються полісахариди — целюлоза і крохмаль. З проміжних продуктів цих перетворень можуть синтезуватися амінокислоти і компоненти ліпідів.
Поиск по сайту: |