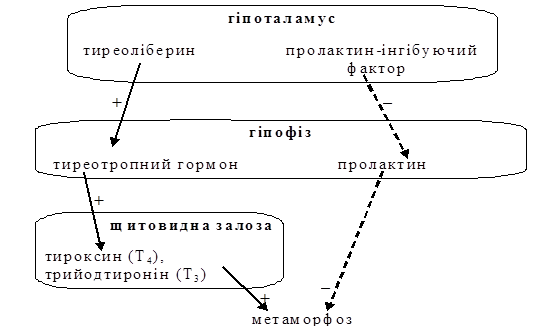|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Метаморфоз та його гормональна регуляціяСтр 1 из 12Следующая ⇒
Змістовний модуль 4. Роль нейроендокринної регуляції в онтогенезі. Молекулярні основи апоптозу та старіння Тема 1. Нейроендокринна регуляція в онтогенезі 1. Загальна характеристика нейроендокринної регуляції. 2. Метаморфоз та його гормональна регуляція. 3. Нейроендокринна регуляція линьки хордових. 4. Гормональна регуляція росту. 5. Механізми гуморальної регуляції в організмі.
Питання 1. Загальна характеристика нейроендокринної регуляції Кожен організм, одноклітинний чи багатоклітинний, є єдиним цілим. Всі його органи тісно пов'язані одне з одним й управляються загальним, точним, злагодженим механізмом. В організмі є так звана гуморальна регуляція і координація роботи окремих органів структури та фізіологічних систем. Вона здійснюється за допомогою особливих високоактивних хімічних речовин, які накопичуються у крові та тканинах у процесі життєдіяльності організму. Механізми регуляції фізіологічних функцій традиційно поділяють на нервові і гуморальні, хоч насправді вони утворюють єдину регуляторну систему, що забезпечує підтримку гомеостазу і пристосувальну діяльність організму. Ці механізми мають численні зв'язки на рівні функціонування нервових центрів, і під час передачі сигнальної інформації еффекторним структурам. Варто сказати, що у здійсненні найпростішого рефлексу як елементарного механізму нервових регуляцій передача сигналів з однієї клітини в іншу здійснюється за допомогою гуморальних чинників -нейромедіаторів. Чутливість сенсорних рецепторів до дії подразників і функціональний стан нейронів змінюється під впливом гормонів, нейромедиаторів, інших біологічно активних речовин, і навіть найпростіших метаболітів і мінеральних іонів (До+,Na+,Ca-+, С1~). Натомість, нервова система може запускати чи виконувати корекцію гуморальних регуляцій. На ранніх етапах еволюції, коли нервова система відсутня, взаємозв'язок між окремими клітинами і навіть органами здійснюється гуморальним шляхом. Але в міру розвитку нервового апарату, у його вдосконалення на найвищих щаблях фізіологічного розвитку гуморальна система дедалі більше підпорядковується нервовій. Гуморальні регуляції здійснюються шляхом поширення сигнальних молекул в рідинах організму за принципом "всім, всім, всім", чи принципу "радіозв'язку". Нервові регуляції здійснюються за принципу "лист із адресою", чи "телеграфної зв'язку". Сигналізація передається від нервових центрів до певних структур, наприклад до певних м'язових волокон чи його групам у певному м'язі. Гуморальние регуляції, зазвичай, здійснюються повільніше, ніж нервові. Швидкість проведення сигналу (потенціалу дії) в швидких нервових волокнах сягає 120 м/с, тоді як швидкість транспорту сигнальною молекули із течією крові в артеріях приблизно 200 раз, а капілярах - в тисячі разів менше. Гуморальні механізми мають значення в регуляції процесів обміну речовин, швидкості розподілу клітин, розвитку і спеціалізації тканин, статевого дозрівання, адаптації зміни умов довкілля. Нервова система у здоровому організмі впливає попри всі гуморальні регуляції, здійснює їх корекцію. Проте у нервової системи є специфічні функції. Вона регулює життєві процеси, потребують швидких реакцій, забезпечує сприйняття сигналів, які приходять від сенсорних рецепторів органів чуттів, шкіри внутрішніх органів. Регулюють тонус та скорочення кісткових м'язів, що забезпечують підтримку пози, переміщення тіла у просторі. Нервова система забезпечує прояв таких психічних функцій, як відчуття, емоції, мотивації, пам'ять, мислення, свідомість, регулює поведінкові реакції, створені задля досягнення корисного пристосувального результату. Питання 2. Метаморфоз та його гормональна регуляція Серед безхребетних непрямий розвиток найкраще вивчений у комах. У них виділяють два основних типи метаморфозу: 1) з повним перетворенням; 2) з неповним перетворенням. При неповному перетворенні (геміметаболії) комаха проходить такі стадії розвитку: яйце, личинка, імаго (мал. 2). З яйця виходить личинка, зовні доволі схожа на дорослу особину. Вона має фасеткові очі, такий самий як і в дорослих ротовий апарат, а на старших стадіях – зовнішні зачатки крил. Спосіб життя личинок і дорослих комах у багатьох видів з неповним перетворенням однаковий. Ці личинки звуться німфами. Окрему модифікацію становлять так звані наяди – личинки одноденок, бабок і веснянок. Вони теж схожі на імаго, мають фасеткові очі й зачатки крил (на старших стадіях ), але живуть у воді й мають спеціальні личинкові органи – зябра, маску (личинки бабок) тощо. Ріст і розвиток личинки супроводжується періодичними линяннями, кількість яких, як правило, стала для певних таксонів (найчастіше, родин). З кожним линянням личинка за розмірами, пропорціями тіла, розвитком крил наближається до дорослої стадії і під час останньоі линьки перетворюється на імаго. При повному перетворенні (голометаболії) комахи проходять такі стадії: яйце, личинка, лялечка, імаго (мал. 3). З яйця виходить личинка, яка різко відрізняється від імаго більшою гомономністю метамерії (тобто, всі сегменти тіла подібні за будовою один до одного), відсутністю зовнішніх зачатків крил, слабшим розвитком органів чуття. Вона не має фасеткових очей, антени її значно коротші; часто ротовий апарат у неї інший, ніж у дорослої комахи (наприклад, у гусені метеликів – гризучий, а в імаго – сисний). Личинки комах із повним перетворенням часто живуть в інших умовах, ніж дорослі. Більшість органів личинки має тимчасовий характер, наприклад черевні ноги в личинок метеликів та пильщиків, шовковидільні залози у гусені метеликів. Личинкові ознаки зберігаються під час усієї личинкової фази. Із кожним линянням личинка лише збільшується в розмірах, а необхідні для метаморфозу зміни відсуваються до останнього личинкового линяння. Закінчивши свій ріст, личинка останньої стадії припиняє живлення, стає нерухомою, линяє востаннє й перетворюється на лялечку. Лялечка нерухома, проте всередині неї відбуваються інтенсивні процеси внутрішньої перебудови – гістоліз та гістогенез. Оскільки характер руху імаго (особливо, політ) і його живлення інакші, ніж у личинкової фази, то м’язи, травна система, ротовий апарат, кінцівки та інші личинкові органи мають повністю перебудуватися. Більшість органів личинки руйнується; цей процес зветься гістолізом. Він відбувається шляхом фагоцитозу, аутолізу (самоперетравлення), а також,– за допомогою літичних ферментів. У різних комах переважає той чи інший механізм, але найчастіше діють усі три: гемоцити виділяють у гемолімфу ферменти, які руйнують тканини; їх залишки фагоцитуються гемоцитами. Крім того, в деяких тканинах відбувається аутоліз. Коли гістоліз досягає свого піку, внутрішні органи лялечки перетворюються на напіврідку масу, яка складається з гемолімфи, збагаченої продуктами розпаду. Не руйнуються лише нервова, статева системи та спинна кровоносна судина. Нервова система може доповнюватися новими клітинами, в ній можуть концентруватися ганглії, проте вона ніколи не втрачає своєї цілісності. Метаморфоз комах відбувається під гормональним контролем з боку нейроендокринної системи (мал. 4). Метаморфоз регулюється нейроендокринним комплексом органів, до якого входять нейросекреторні клітини мозку, кардіальні тіла, прилеглі тіла та парна проторакальна (передньогрудна) залоза. Нейросекреторні клітини мозку виробляють проторакотропний гормон. Через довгі аксони цих клітин він надходить до кардіальних тіл, а з них – у гемолімфу. У личинок проторакотропний гормон стимулює роботу проторакальних залоз, які починають виділяти гормон линьки – екдізон. Прилеглі тіла виділяють ювенільний гормон. Концентрація екдізону мала в періоди між линьками і висока – в період линьки; а концентрація ювенільного гормона висока протягом всієї личинкової стадії, і знижується перед линянням на імаго чи лялечку. Екдізон в периферичних тканинах перетворюється на свою активну форму – екдістерон і впливає на клітини гіподерми, спричиняючи спочатку синтез ферментів, які розчиняють стару кутикулу, а пізніше – синтез матеріалу для побудови нової кутикули. Крім того, екдізон стимулює ріст і диференціацію всіх тканин личинки, в тому числі,– й клітин імагінальних дисків. Але при високій концентрації ювенільного гормону екдізон не може активувати розвиток імагінальних дисків і тому стимулює лише секрецію личинкової кутикули, і линяння завершується появою чергової личинкової стадії. Зниження концентрації ювенільного гормону веде до зняття блокуючого впливу на імагінальні диски, екдізон стимулює їх ріст та диференціацію, і відбувається линяння личинки на лялечку (або останню личинкову стадію при неповному метаморфозі).
Мал. 4. Гормональна регуляція метаморфозу у комах. Метаморфоз у амфібійрегулюється гормонами гіпоталамо-гіпофізарно-тиреоїдної системи та пролактином (мал. 6). Гормони щитовидної залози (трийотиронін і тироксин) стимулюють метаморфоз. Пролактин є свого роду личинковим гормоном, який стимулює личинковий ріст і гальмує метаморфоз. Гіпоталамус виробляє тиреоліберин, який стимулює утворення тиреотропного гормона в гіпофізі. Тиреотропний гормон стимулює утворення трийодтироніну та тироксину щитовидною залозою. В гіпоталамусі також виробляється пролактин-інгібуючий фактор, який гальмує утворення пролактину гіпофізом (мал. 6).
Рис. 6. Гормональна регуляція метаморфозу в амфібій У пуголовків гіпоталамус недорозвинутий. Тому він виробляє мало тиреоліберину, що веде до виділення малої кількості тиреотропного гормона, і, як наслідок, до утворення малої кількості тироксину і трийодтироніну. Внаслідок тої ж таки недорозвинутості гіпоталамуса, він виділяє мало пролактин-інгібуючого фактора, що веде до утворення великої кількості пролактину. Тому стимулюється личинковий ріст і гальмується метаморфоз. З розвитком гіпоталамуса він починає виробляти все більше тиреоліберину і пролактин-інгібуючого фактора, що веде до утворення все більшої кількості тироксину і трийодтироніну і все меншої кількості пролактину. В кінці кінців концентрація пролактину падає нижче певного порогового рівня, а концентрація гормонів щитовидної залози піднімається вище певного порогового рівня, що призводить до запуску процесів метаморфозу. Тиреоїдні гормони впливають на клітини-мішені на рівні транскрипції (тобто, впливають безпосередньо на синтез іРНК). Реакція тканин на дію цих гормонів спеціфічна: одні гинуть, дегенерують, інші – не реагують, ще одні – активуються. Така специфічність є властивістю реагуючої тканини. В ході метаморфозу повинна бути забезпечена певна послідовність змін (наприклад, розвиток кінцівок повинен відбутися раніше, ніж редукція звоста, а не навпаки); ймовірно, що це досягається за рахунок того, що різні тканини реагують на різні порогові концентрації тиреоїдних гормонів. Питання 3.
Поиск по сайту: |