|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Специфичность действия ферментов
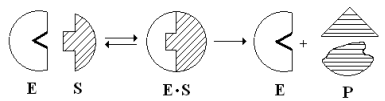
Понятие Е·S комплекса и АЦФ тесно взаимосвязаны с особым свойством ферментов – их специфичностью. По степени специфичности (в порядке ее снижения) различают: I. Стереохимическую субстратную специфичность – в этом случае ферменты катализируют только 1 форму S (1 изомер). Например, фумаратгидратаза катализирует только превращение фумаровой кислоты, но не катализирует превращение ее изомера – малеиновую кислоту. II.Абсолютную субстратную специфичность – Е превращает только 1S. Например, уреаза превращает только мочевину. III. Абсолютная групповая S-ную специфичность. Ферменты действуют на группу сходных S-в. Например, алкоголь ДГ превращает не только этанол, но и другие алифатические спирты. IV. Относительную групповую S-ную специфичность. Фермент воздействует не на группу молекул S, а на определенные связи определенных групп S-в. Например, пепсин и трипсин специфичны по отношению к пептидным связям в различных белках. V. Относительную S-ную специфичность. Фермент катализирует, превращаясь в S-в, относящимся к различным группам химических соединений. Например, фермент цитохром-450 катализирует реакции гидроксилирования до 7000 разных S-в. Это наименее специфичная ферментная система.
Существует две теории объяснения специфичности ферментов. Гипотеза Э. Фишера – гипотеза «ключа и замка» или гипотеза «шаблона». По Фишеру, фермент – это жесткая структура, АЦФ которого - точный «слепок» S-та. Если S подходит к Е как ключ к замку, то реакция произойдет. Если же S немного изменен («ключ»), то он не соответствует АЦФ («замку»), и реакция становится невозможной. Несмотря на логичность такого объяснения, гипотеза Фишера не объясняет, на чем тогда основаны абсолютная и относительная групповая специфичность. Например, цитохром-450 соединяется с таким большим количеством S-в, различных по строению. Эти внешние противоречия объясняет гипотеза Кошленда, или гипотеза вынужденного соответствия. По мнению Кошленда, молекула фермента не «жесткая», а гибкая структура и конфигурация фермента и его АЦФ начинают изменяться в момент присоединения фермента к S или другим лигандам. При образовании Е-S комплекса кроме геометрической комплементарности имеет место и электростатическая, которая осуществляется благодаря спариванию противоположно заряженных молекул Е и S. В действительности, видимо, имеют место оба варианта присоединения.
Гипотеза Кошленда позволяет объяснить, почему происходит превращение близких аналогов S-в. Если «ложный» субстрат (квази-S) отличается от природного и АЦФ принимает конформацию близкую к истинному субстрату, то расстановка каталитических групп в таком Е-S комплексе позволит осуществить реакцию. Этот «обман» фермент как бы не замечает, хотя реакция идет и не так быстро, как с истинным субстратом. Если конфигурация квази субстрата не позволяет правильно расположиться каталитической группе, то в этом случае реакция не пойдет. Т.е. если диапазон конформационных перестроек ограничен до одной единственно возможной, то фермент высокоспецифичен, а если возможности перестройки АЦФ велики, то фермент срабатывает и на квази субстраты. Зависимость Vфр от рН-среды
При оптимальном значении рН функциональные группы АЦФ и сам S находятся в наиболее предпочтительной для связи форме. Некоторые ферменты имеют оптимальную рН, резко отличающиеся от физиологических значений, пепсин на 100% активен при рН = 1,5-2,5; аргиназа – при рН = 10. Зависимость Vфр от температуры
Для некоторых ферментов денатурации наступает при 0ºС. Для любых химических реакций при повышении температуры на каждые 10ºС V реакции увеличивается в 2-3 раза, для ферментативных реакций этот коэффициент ниже – 2 и даже меньше. Исключение: термостабильный фермент адениматциклаза выдерживает температуру 100ºС, а фермент каталаза активен при 0ºС. Зависимость Vфр от конц. S. Механизм действия ферментов описывается уравнением Михаэлиса-Ментен. Установить зависимость Vфр от [S] можно графически.
б)методом обратных величин Лайнцивера-Берка, где зависимостьVфр от [S] рассчитывают в обратных величинах.
Поиск по сайту: |



 в растворе избыток Е (3 мол S, 5 мол Е) это участок насыщения фермента S-том.
в растворе избыток Е (3 мол S, 5 мол Е) это участок насыщения фермента S-том.