|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
ДИССИПАТИВНЫЕ СТРУКТУРЫ
Единый процесс развития охватывает явления живой и неживой природы и общества, поэтому естественно описать весь процесс развития на одном языке, в рамках единой схемы, с использованием общей терминологии. В эволюционной теории для описания процессов развития используют триаду: изменчивость, наследственность, отбор. Эти же факторы рассматривает Н.Н.Моисеев в своей монографии «Алгоритмы развития». Изменчивостью он называет любые проявления стохастичности и неопределенности. Наследственностью – способность материи сохранять свои особенности, способность изменяться от прошлого к будущему, способность будущего зависеть от прошлого. Принципами отбора он называет те принципы, которые вызывают к существованию более или менее устойчивые образования, ими являются законы сохранения, законы физики и химии в частности, второй закон термодинамики, вариационные принципы и т.д.. Задача состоит в том, чтобы выявить то общее содержание, которое присуще любым процессам развития. Моисеев строит классификацию принципов отбора и рассматривает с единой точки зрения его механизмы. Он выделяет два разных класса отбора. Это адаптационные механизмы (определение множества состояний системы, которые будут обеспечивать ее устойчивость при данных условиях внешней среды) и бифуркационные механизмы (качественный скачок, изменение организации системы). Определяющую роль эволюционного периода развития систем играют такие понятия, как адаптация, устойчивость, стабилизирующий отбор. Необходимым условием существования живых организмов является постоянство внутренней среды. Гомеостазис (от греческого «гомео» – тот же, «стазис» – состояние) рассматривается биологами как способность биологических систем противостоять изменениям внешней среды и сохранять состояние равновесия. Например, только благодаря механизмам поддержания гомеостазиса некоторые растения могут жить на ядовитых отвалах рудников. Есть растения – концентраторы металлов – алюминия, молибдена, никеля, свинца, стронция. При этом для предотвращения отравления тканей в растениях синтезируются специфические белки (определяющие устойчивость к высоким концентрациям металлов), изменяются количество и качество корневых выделений, тяжелые металлы связываются в клетках дубильными веществами и органическими кислотами. Механизмы поддержания гомеостазиса исторически закреплены и направлены на повышение устойчивости организма в онтогенезе, что обеспечивает успех в воспроизведении потомства. Развитие – это борьба двух противоположных тенденций – сохранение гомеостазиса и поиск новых организационных форм, уменьшающих локальную энтропию. Этапность развития органического мира, на которой в значительной мере базируется периодизация геологической истории, – твердо установленный факт. Однако общая теория этапности разработана еще очень слабо, особенно это касается наиболее принципиального ее раздела «О закономерностях перехода от одного этапа к другому». До сих пор обычно недооцениваются изменения биоценотических связей в экосистемах прошлого, вызывавшиеся появлением новых групп организмов. Вместе с тем чрезвычайно большое внимание уделяется возможной роли глобальных катастроф. Согласно концепции прерывистого равновесия, разработанной американскими исследователями С.Гоулдом, Н.Элдриджем и С.Стэнли, эволюция, во всяком случае на видовом уровне, по крайней мере в 95% случаев идет не непрерывно, а своего рода скачками. Предполагается, что виды остаются практически неизменными на протяжении буквально миллионов лет, а затем за несколько десятков или сотен лет происходит формирование новых видов. Переход от вида к виду совершается в ее свете не посредством скачка в одном поколении, а путем накопления мутации и отбора. Всякое значительное изменение экологических условий влечет за собой перестройку всей организации сообщества животных. Например, у животных, ведущих одинокий образ жизни, в случае необходимости возникает строгая иерархическая структура. В период нехватки корма такая организация сообщества определяет очередность доступа к пище. Доминирование может не только разделять, но и объединять животных, оно способствует процессу локализации, образованию структуры в сообществе взамен агрессивных взаимодействий особей. Биологические системы обладают способностью сохранять и передавать информацию в виде структур и функций, возникших в прошлом в результате длительной эволюции. Открыты подвижные генетические элементы, которые оказались замешаны в таких общебиологических явлениях, как азотфиксация, злокачественный рост клеток, работа иммунной системы и приспособление бактерий к антибиотикам, нестабильные мутации, материнская наследственность. Нестойкое, нестабильное состояние гена, когда он начинает мутировать в десятки, сотни раз чаще обычного, связано не с изменениями внутри самого гена, а с введением в район его расположения определенного «контролирующего» элемента, способного блуждать по хромосомам. Эти элементы влияют на «включение» и «выключение» генов, т.е. на темп наследственной изменчивости. Одно из самых удивительных открытий для генетиков в последние 15-20 лет состояло в осознании повсеместности подвижных элементов, общности их строения и причастности к самым разным генетическим явлениям. Подвижные гены имеют на одном и другом конце повторы. Такие генетические тексты, обрамленные повторами, начинают вести свою отдельную от общей наследственной системы жизнь. Именно такого рода структуры получают возможность увеличивать число своих копий в хромосомах. Они подчиняют своему звучанию близлежащие гены, которые либо замолкают, либо усиливают активность, либо начинают работать в другом режиме. Включив в свой состав участок ДНК, отвечающий за самоудвоение, подвижный элемент превращается в плазмиду, которая самостоятельно размножается вне дочерней хромосомы у бактерий и вне ядра в клетках высших организмов. В классической генетике: мутация возникает случайно; им подвержены единичные особи; их частота очень мала. В «подвижной генетике» изменения не случайны, зависят от типа подвижного элемента; им подвержены много особей; их частота велика, может достигать десятка процентов. Именно с мобильностью активных элементов связывают обнаруженные в природных популяциях дрозофил регулярные вспышки мутации определенных генов. Темп мутационного процесса непостоянный, так, время от времени популяции или виды вступают в «мутационный» период. Самое поразительное открытие в генетике за последнее время – это возможность с помощью мобильных элементов переносить гены или группы генов от одних видов к другим (иногда к самым далеким), т.е. благодаря перемещающимся элементам генофонды всех организмов объединены в общий генофонд всего живого мира. Это особенно ярко продемонстрировали плазмиды с детерминантами устойчивости к антибиотикам в колоссальном эксперименте, невольно поставленном человеком на бактериях. С помощью генсектицидов человек расширяет эксперимент на насекомых, и в ответ их популяции, вероятно, охватываются определенными, быстро распространяющимися генетическими элементами, повышающими устойчивость организма («генетическая экспансия»). Предполагается, что когда-то в клетках насекомых поселились бактерии – симбионты, которые постепенно передали большинство своих генов в ядро и превратились в митохондрии и пластиды. Это замечательный пример переноса генов от про– к эукариотам. Способность клеток одного вида воспринимать ДНК от других, иногда эволюционно далеких видов, возможность горизонтального переноса генов считается «одним из главных чудес XX века». Классическая генетика гласит: каждый ген располагается на своей хромосоме и занимает на ней строго фиксированное положение. Сейчас известно много вариантов перемещающихся элементов, которые могут менять свое место на хромосоме и даже перемещаться с хромосомы на хромосому. Таким образом могут рождаться новые признаки организма. Однако способность системы обмениваться информацией с внешней средой, увеличивать или уменьшать число элементов-признаков, сохранять устойчивость еще не делает эту систему развивающейся. Порождаемая неравновесными внешними условиями неустойчивость приводит к увеличению интенсивности диссипации, вследствие чего создаются условия возникновения новой неустойчивости. Иными словами, в системе увеличивается интенсивность протекания некоторых необратимых процессов, благодаря чему и отклонение системы от равновесия становится еще большим. Это означает, что вероятность существования такого класса флуктуаций, по отношению к которым новые процессы становятся неустойчивыми, возрастает. С другой стороны, если бы в результате возникновения неустойчивости интенсивность диссипации снижалась, то система по своим свойствам приблизилась бы к некоторой равновесной замкнутой системе, т.е. к состоянию, в котором затухают любые флуктуации. Такой механизм можно изобразить следующим образом (И.Р.Пригожин, И.Стенгерс): Интенсивность диссипации, т.е. увеличение энтропии, можно связать с интенсивностью роста числа новых элементов-признаков в системе. Если флуктуации вызывают интенсивный рост новых элементов и между ними не успевают образовываться связи, организация системы нарушается, энтропия возрастает, система становится структурно неустойчивой. Существование неустойчивости можно рассматривать как результат флуктуации, которая сначала была локализована в малой части системы, а затем распространилась и привела к новому макроскопическому состоянию. Исследования школы Пригожина показали, что понятия структурной устойчивости и порядка через флуктуации применимы к системам различной природы, в том числе экономическим, социальным: «Пределов для структурной устойчивости не существует. Неустойчивости могут возникать в любой системе, стоит лишь ввести подходящие возмущения. Мутации и «новорожденные» элементы возникают стохастически и собираются в единую систему господствующими в данный момент детерминистическими принципами. Это позволяет нам надеяться на непрестанную генерацию «новых типов» и «новых идей», которые могут быть включены в структуру системы, обеспечивая тем самым ее непрерывное развитие». Удаленность от равновесия, нелинейность может служить причиной возникновение упорядоченности в системе. Биологическая упорядоченность, генерация когерентного света лазером, возникновения пространственной и временной упорядоченности в химических реакциях и гидродинамике, автоволны в различных средах, наконец, функционирование экосистем в животном мире или жизнь человеческого общества – все эти примеры являются поразительной иллюстрацией явлений самоорганизации, образования диссипативных структур. Эти структуры наряду с замечательными регуляторными свойствами проявляют необычайную гибкость и разнообразие. Как показали работы школы Пригожина, важнейшей общей чертой широкого класса процессов самоорганизации является потеря устойчивости и последующий переход к устойчивым диссипативным структурам. В точке изменения устойчивости в результате ветвления должны возникнуть по меньшей мере два решения, соответствующие устойчивому, близкому к равновесному состоянию и диссипативной структуре. Для диссипативных структур характерна устойчивость, которая одновременно является структурной и функциональной. Эволюцию можно рассматривать как проблему структурной устойчивости. Система не всегда является структурно устойчивой, причем эволюция диссипативной структуры определяется последовательностью событий в соответствии со схемой (Г.Николис, И.Р.Пригожин). Шмальгаузен в качестве одного из факторов эволюции выделял борьбу за существование. Согласно теории конкуренции, близкие виды могут населять район лишь в том случае, если они эффективно делят между собой необходимые ресурсы. Хатгинсон провозгласил принципиально новую программу изучения неравновесных сообществ, чьи принципы организации коренным образом отличаются от тех, к которым привыкли экологи, рассматривающие сообщество как жестко организованную совокупность видов. В альтернативном подходе прямая конкуренция за пищу подменяется сложными и динамичными информационными связями. В зависимости от условий метаболиты (продукты жизнедеятельности) данного вида либо угнетают, либо, наоборот, стимулируют размножение других видов. При этом возможны как негативные, так и позитивные межвидовые отношения. Разные виды не только не стремятся окончательно вытеснить друг друга, но как бы «удерживают» в сообществе те виды, численность которых падает ниже определенного уровня. В теории конкуренции выражена концепция детерминизма; конкуренция определяет численность, облик и эволюцию существующих видов, потребляющих одну и ту же пищу. Другая парадигма: близкие виды, расположенные в экосистеме на одном трофическом уровне, всегда живут среди избытка пищи, поскольку их численность эффективно ограничивается сложным комплексом причин, в том числе и конкуренцией. Согласно этой парадигме биологическое сообщество можно рассматривать как диссипативную структуру, которая, находясь в неравновесном состоянии, постоянно ведет обмен с внешней средой. В исходной неупорядоченной системе за счет последовательно реализующихся неравновесных неустойчивостей, за счет когерентного поведения элементов может возникнуть функциональная организованность. Приведем пример из нового быстро развивающегося научного направления – адаптологии. Клод Бернар назвал приспособление адаптационно-итоговой проблемой всей физиологии. Адаптация человека и животных представляет собой процесс, в течение которого организм приобретает отсутствовавшую ранее устойчивость к определенным факторам среды и в результате решает задачи, ранее несовместимые с жизнью. При всем разнообразии приспособительных процессов в них есть сходство. На первом этапе адаптации к любому новому фактору организм подходит к максимуму своих возможностей, к критической (бифуркационной) точке. Если человек или животное не погибает, а фактор по-прежнему действует, то возможности живой системы возрастают, и на смену аварийной стадии в большинстве случаев приходит стадия эффективной и устойчивой привычки (возникает диссипативная структура). Согласно учению Ухтомского, формирование систем, обеспечивающих адаптацию, происходит следующим образом. Под влиянием внешних раздражителей в организме формируются, сменяя друг друга, созвездия возбужденных нервных центров – доминанты. Именно доминанты обеспечивают приспособление организма к среде, его выживание, а если надо – и преобразующее влияние на среду. В доминирующей системе образуется структура, которая увеличивает мощность системы и становится материальной основой долговременной адаптации (например, после перенесенных заболеваний сердца – рост сердца в целом, соотношение структур в клетках сердца меняется так, что орган в целом становится более мощным и эффективным). Стресс – реакция (флуктуация) нужна для того, чтобы возникла доминирующая в адаптации система и образовался структурный след (новая структура, обеспечивающая новый способ функционирования). Доминирующая система работает под нагрузкой, в ее клетках возрастает производство нуклеиновых кислот и белков, строятся новые структуры. Смотря по тому, к какому фактору приспосабливается организм, доминирующая система и ее след выглядят по-разному. Но этот след всегда увеличивает мощность системы, материально обеспечивает переход к долговременной адаптации. Как и любая другая архитектура, архитектура системного структурного следа есть отражение задачи, которую среда выдвигает перед организмом. Перед современным человеком чаще возникают задачи, требующие не физической, а умственной адаптации. При обучении – формировании условных рефлексов, навыков, идей возбуждение нейронов в высших отделах мозга активизирует в них производство нуклеиновых кислот и белков. Белки из тела нейронов поступают в отростки, которыми нервные клетки контактируют между собой. Так, перестраиваются межнейронные связи, возникают новые нейронные сети. Сегодня модели нейронных и иммунных сетей строятся в рамках представлений о диссипативных структурах. Переход системы после критической точки из неустойчивого состояния к устойчивому (к диссипативной структуре) можно рассматривать как качественный скачок в развитии системы, в результате которого возросла организованность, упорядоченность системы, Место нервной клетки в структуре мозга определяется очень рано – у млекопитающих и человека в основном до рождения, и это место далеко не случайно. Это не значит, что структура мозга генетически полностью предопределена. Связи формируются преимущественно после рождения, а следовательно, в различной мере зависимы от внешней среды. Некоторые связи будут обязательно, другие могут быть, а могут и не быть, третьи могут оказаться патологическими. Поэтому нормальное развитие мозга зависит от гармонии усложнения внутренней и внешней среды организма. При этом для человека наиболее существенным фактором внешней среды, гармонизирующим внутреннюю структуру мозга, являются другие люди. Работы нейрофизиологов в области исследования пространственной организации как отдельных образований, так и целых систем мозга показали, что структурно-функциональной единицей мозга служит не отдельный нейрон, а популяция нейронов, пространственно организованная в виде вертикальной колонки, расположенная в одном или нескольких слоях коры. Эволюционные преобразования мозга в определяющей мере зависят от изменений взаиморасположения элементов, т.е. от изменений конструкций, как отдельных структур мозга, так и мозга в целом. И в меньшей степени – от изменений самих элементов. Выявлена зависимость между увеличением вертикальной упорядоченности и усложнением функций мозга. Максимальная величина упорядоченности соответствовала наиболее новым и сложноорганизованным формациям коры мозга человека, имеющим отношение к восприятию речи, ее воспроизводству.
Законы Менделя
В 1856–66 годах чешским монахом Грегором Менделем были поставлены знаменитые опыты, результатом которых стало появление новой науки – генетики. Объектом для экспериментов был выбран огородный горох, так как существует множество его сортов, чётко различающихся по ряду признаков; растения легко выращивать и скрещивать. Успех Менделя объясняется тщательным планированием и аккуратным проведением экспериментов, а также наличие большого количества опытов, позволявших получить статистически достоверные сведения.
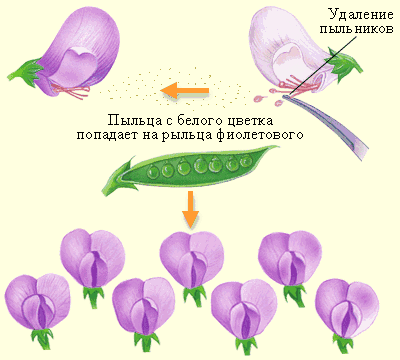
Для своих первых опытов Мендель выбирал растения, чётко различающиеся по какой-либо паре признаков, например, по расположению цветов («пазушные» или «верхушечные»). Выращивая растения каждого типа на протяжении нескольких поколений, Мендель убедился в их пригодности для проведения эксперимента. Мендель проводил скрещивание – опылял растения одного типа пыльцой растений другого типа. Ряд предосторожностей (например, удаление тычинок у цветков, которые впоследствии опылялись, и надевание колпачков на цветы, чтобы избежать дополнительного опыления со стороны других растений) позволили получить достоверные результаты. Во всех случаях из семян, собранных с этих гибридов, вырастали растения с пазушными цветками. Признак «пазушные цветки», наблюдаемый у гибридов первого поколения, был назван доминантным, признак «верхушечные цветки» – рецессивным. Далее растениям первого гибридного поколения была предоставлена возможность самоопылиться. Во втором гибридном поколении у части растений образовались пазушные цветки, а у другой части – верхушечные. Мендель предположил, что признак «верхушечные цветки» присутствовал и в первом поколении, но в скрытом виде. Во всех подобных опытах, проведённых с какой-либо парой признаков, примерно три четверти гибридов второго поколения обладали признаком, проявлявшимся и в первом поколении гибридов (его назвали доминантным), а четверть потомства второго поколения обладала признаком, не проявившимся у гибридов первого поколения (рецессивным). Важно, что чем больше опытов было поставлено, тем ближе был полученный результат к отношению 3 : 1. На основании этой серии опытов были сделаны следующие выводы: - У родительских растений было по два одинаковых «фактора» (например, «пазушные цветки» либо «верхушечные цветки»). - Гибриды первого поколения получили по одному фактору от каждого родителя, причём эти факторы не слились, а сохранили свою индивидуальность. Таким образом, был сформулирован закон расщепления (первый закон Менделя).
Итак, каждый признак организма контролируется парой вариантов гена (или, как говорят, порой аллелей). Если в генотипе организма имеются аллели обоих типов, то один из них (доминантный) будет проявляться, полностью подавляя другой (рецессивный). При мейозе каждая пара аллелей расщепляется, и с каждой гаметой как дискретная, не изменяющаяся величина, может передаваться только один аллель. Передача генов потомкам находится в полном соответствии с теорией вероятности. Вероятность того, что гамета, полученная от гибрида первого поколения, будет нести доминантный аллель, равна 1/2. Вероятность каждой из четырёх комбинаций при оплодотворении составит 1/4; из них три комбинации будут содержать доминантный аллель и приведут к появлению особей с доминантным признаком. Первая из этих комбинаций содержит исключительно доминантные аллели – AA (говорят, что она гомозиготна по доминантному аллелю), а две другие содержат по одному доминантному и одному рецессивному аллелю – Aa (гетерозиготны). В четвёртой комбинации будут содержаться только рецессивные аллели; они будут соответствовать потомству с рецессивным признаком (то есть будут гомозиготны по рецессивному аллелю). Гомозиготные особи при последующем самоопылении не расщепляются (дают единобразное потомство). В потомстве самоопыляющихся гетерозиготных особей наблюдается расщепление по внешним признакам в том же соотношении 3 : 1.
Ген обычно обозначается первой буквой, с которой начинается название доминантного аллеля этого гена (например, A). При этом доминантный аллель обозначается прописной буквой (A), а рецессивный – строчной (a). Гибрид первого поколения в описанных опытах гетерозиготен по своему генотипу, но обладает доминантным фенотипом (то есть имеет доминантный признак). Во втором поколении особи с доминантным фенотипом могут обладать как гомозиготным, так и гетерозиготным генотипом. Чтобы выяснить генотип гибрида второго поколения за одно скрещивание, необходимо произвести возвратное (анализирующее) скрещивание с особью, гомозиготной по рецессивному аллелю изучаемого гена. Если у всех потомков от этого скрещивания проявится доминантный фенотип, то особь с определяемым генотипом была гомозиготна по доминантному признаку. Если же появятся особи как с доминантными, так и рецессивными признаками (в примерном соотношении 1:1), то изучаемая особь была гетерозиготна.
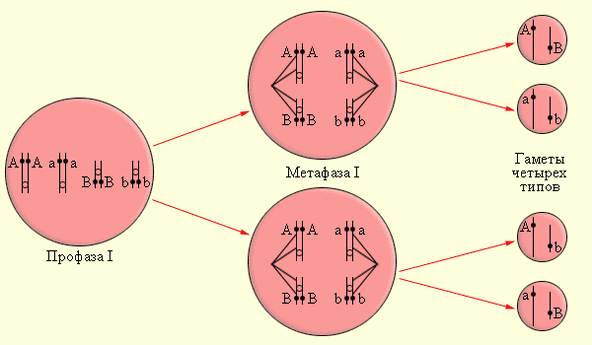
В описанных опытах проводилось моногибридное скрещивание – брались особи, различавшиеся только по одному признаку. В дальнейшем Мендель перешёл к изучению дигибридного скрещивания, когда по той же методике ставились опыты над чистосортными (гомозиготными) особями, различающимися по двум признакам (например, жёлтые и зелёные семена, морщинистые и гладкие семена). В результате, во втором поколении могли получиться особи с семенами четырёх типов: жёлтые и гладкие, жёлтые и морщинистые, зелёные и гладкие, зелёные и морщинистые. Соотношение разных фенотипов во втором поколении составило примерно 9 : 3 : 3 : 1. При этом для каждой пары признаков приближённо выполнялось соотношение 3 : 1. На основании этого Мендель вывелпринцип независимого распределения (второй закон Менделя).
Схему дигибридного скрещивания удобно записывать в специальной таблице – так называемой решётке Пеннета; при этом количество возможных ошибок при определении генотипа потомства сводится к минимуму. Все генотипы мужских гамет вносятся в заголовки вертикальных столбцов, а все генотипы женских гамет – в заголовки горизонтальных. Если вернуться к примеру с семенами гороха, то можно выяснить, что вероятность появления во втором поколении особей с гладкими семенами (доминантный аллель) равняется 3/4, с морщинистыми семенами – 1/4 (рецессивный аллель), с жёлтыми семенами – 3/4 (доминантный аллель) и с зелёными семенами – 1/4 (рецессивный аллель). Таким образом, вероятности сочетания аллелей в генотипе равны: - гладкие и жёлтые – 9/16 (3/4 ∙ 3/4); - гладкие и зелёные – 3/16 (3/4 ∙ 1/4); - морщинистые и жёлтые – 3/16 (1/4 ∙ 3/4); - морщинистые и зелёные – 1/16 (1/4 ∙ 1/4);
Законы Менделя не были восприняты мировым научным сообществом. В 1900 году Хуго де Фриз, Карл Корренс и Эрих Чермак независимо друг от друга заново открыли законы Менделя, сформулировав их в форме, близкой к современной. Одновременно по мере совершенствования микроскопа стала очевидной роль ядра и хромосом в передаче наследственных факторов. В результате была создана хромосомная теория наследственности, согласно которой каждая пара генов локализована в паре хромосом, причём каждая хромосома несёт по одному фактору. Последние исследования показали, что наследственные признаки могут передаваться не только в хромосомах, но и через цитоплазму (будучи локализованными в генетическом материале митохондрий и пластид). Цитоплазматическая наследственность передаётся только по материнской линии (при оплодотворении митохондрии и пластиды из мужских половых клеток не попадают в зиготу).
Поиск по сайту: |