|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Образование желчных кислот
Синтез желчных кислот идет в эндоплазматическом ретикулуме при участии ци-тохрома Р450, кислорода, НАДФН и аскорбиновой кислоты. 75% ХС образуемого в печени участвует в синтезе желчных кислот. При экспериментальном гиповитами-
нозе С у морских свинок развивались,кроме цинги,атеросклероз и желчнокаменнаяболезнь. Это связано с задержкой ХС в клетках и нарушением растворения его в
желчи. Желчные кислоты (холевая, дезоксихолевая, хенодезоксихолевая) синтези-
руются в виде парных соединений с глицином – гликопроизводные и с таурином – тауропроизводные, в соотношении 3:1 соответственно.
Кишечно-печеночная циркуляция
Это непрерывная секреция желчных кислот в просвет кишечника и их реабсорб-ция в подвздошной кишке. В сутки происходит 6-10 таких циклов. Таким образом, небольшое количество желчных кислот (всего 3-5 г) обеспечивает переваривание
липидов, поступающих в течение суток.
Нарушение желчеобразования
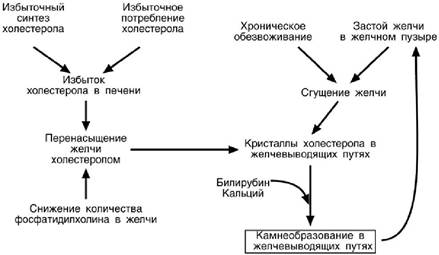
Нарушение желчеобразования чаще всего связаны с хроническим избытком ХС в организме, так как желчь является единственным способом его выведения. В ре-зультате нарушения соотношения между желчными кислотами, фосфатидилхолином и холестерином образуется перенасыщенный раствор холестерина из которого по-следний осаждается в виде желчных камней . Кроме абсолютного избытка ХС в развитии заболевания играет роль недостаток фосфолипидов или желчных кислот при нарушении их синтеза. Застой в желчном пузыре, возникающий при неправиль-ном питании, приводит к сгущению желчи из -за реабсорбции воды через стенку, не-достаток воды в организме также усугубляет эту проблему.
Считается, что желчные камни имеются у 1/3 населения Земли, к пожилому возрасту эти значения достигают 1/2. Интересны данные о способности ультразвукового исследования выявлять
желчные камни только в 30% имеющихся случаев.
Лечение
1. Хенодезоксихолевая кислота в дозе 1 г/сут. Вызывает снижение осаждения ХС
и растворение ХС камней. Камни размером с горошину без билирубиновых наслое-
ний растворяются в течение полугода.
2. Ингибирование ГМГ-S-КоА-редуктазы (ловастатин) – снижает в 2 раза синтез
ХС.
3. Адсорбция ХС в желудочно-кишечном тракте (холестираминовые смолы,
Questran) и предотвращение его всасывания.
4. Подавление функции энтероцитов (неомицин) – снижение всасывания жиров.
5. Хирургическое удаление подвздошной кишки и прекращение реабсорбции
желчных кислот.
Всасывание липидов.
Происходит в верхнем отделе тонкого кишечника в первые 100 см.
1. Короткие жирные кислотывсасываются без каких либо дополнительных ме-ханизмов, напрямую.
2. Другие компоненты образуют мицеллы с гидрофильным и гидрофобным
слоями. Размеры мицелл в 100 раз меньше самых мелких эмульгированных жиро-вых капелек. Через водную фазу мицеллы мигрируют к щеточной каемке слизистой
оболочки.
Относительно самого механизма всасывания липидов нет устоявшегося пред-ставления. Первая точка зрения заключается в том, что мицеллы проникают внутрь
клетки целиком путем диффузии без затраты энергии. В клетках происходит распад
мицелл и выход желчных кислот в кровь, ЖК и МАГ остаются и образуют ТАГ. По другой точке зрения,поглощение мицелл идет пиноцитозом.
И, наконец, в-третьих, возможно проникновение в клетку только липидных ком-
понентов, а желчные кислоты всасываются в подвздошной кишке. В норме всасывается 98% пищевых липидов.
Нарушения переваривания и всасывания могут возникать
• при болезнях печени и желчного пузыря, поджелудочной железы, стенки кишеч-ника,
• повреждении энтероцитов антибиотиками (неомицин, хлортетрациклин);
• избытке кальция и магния в воде и пище, которые образуют соли желчных ки-слот, препятствуя их функции.
Ресинтез липидов
Это синтез липидов в стенке кишечника из посту-
пающих сюда экзогенных жиров, частично могут ис-пользоваться и эндогенные жирные кислоты.
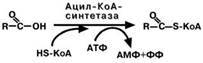
При синтезе триацилглицеролов поступившая
жирная кислота активируется через присоединение ко-
энзима А. Образовавшийся ацил-S-КоА участвует в реакциях синтеза триацилглице-
ридов по двум возможным путям.
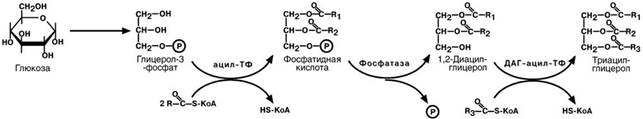
Первый путь–2-моноацилглицеридный,происходит при участии экзогенных2-МАГ и ЖК в гладком эндоплазматическом ретикулуме: мультиферментный комплекс триглицерид-синтазы формирует ТАГ
При отсутствии 2-МАГ и высоком содержании ЖК активируется второй путь,
глицеролфосфатный механизм в шероховатом эндоплазматическом ретикулуме.Источником глицерол-3-фосфата служит окисление глюкозы, так как пищевой глице-
рол быстро покидает энтероциты и уходит в кровь.
Холестерол этерифицируется с использованием ацилS- КоА и фермента АХАТ. Реэтерификация холестерола напрямую влияет на его всасывание в кровь. В настоящее время ищутся возможности подавления этой реакции для снижения концентрации ХС в крови.
Фосфолипидыресинтезируются двумя путями–с использованием1,2-МАГ длясинтеза фосфатидилхолина или фосфатидилэтаноламина, либо через фосфатид-ную кислоту при синтезе фосфатидилинозитола.
Транспорт липидов
Липиды транспортируются в водной фазе крови в составе особых частиц – ли-попротеинов.Поверхность частиц гидрофильна и сформирована белками,фосфо-липидами и свободным холестеролом. Триацилглицеролы и эфиры холестерола со-ставляют гидрофобное ядро.
Белки в липопротеинах обычно называются апобелками,выделяют несколько их типов–А, В, С, D, Е. В каждом классе липопротеинов находятся соответствующие ему апобелки, вы-полняющие структурную, ферментативную и кофакторную функции.
Липопротеины различаются по соотноше-
нию триацилглицеролов, холестерола и его
эфиров, фосфолипидов и как класс сложных белков состоят из четырех классов.
• хиломикроны (ХМ);
• липопротеины очень низкой плотности (ЛПОНП, пре-β-липопротеины, пре-β-ЛП); • липопротеины низкой плотности (ЛПНП, β-липопротеины, β-ЛП);
• липопротеины высокой плотности (ЛПВП, α-липопротеины, α-ЛП).
Поиск по сайту: |