|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Метамерия у простейших, кишечнополостных и низших червей
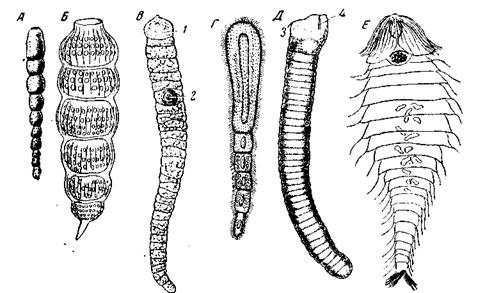
Метамерию мы встречаем во всем животном царстве, начиная с простейших. Среди простейших наиболее распространена метамерия у Fo-raminifera. Здесь она возникает обычно путем неполного деления и выражается весьма явственно в строении раковины, очень часто состоящей из ряда метамерно расположенных камер. Самый простой и наглядный случай представляет раковина Nodosaria или Nodulina (рис. 25, А), все камеры которой бывают обычно расположены вдоль прямолинейной оси.
А – Nodulina nodulosa (Foraminifera) (по Brady); Б – Arthrostrobus articulatus (Radiolaria Nasselaria) (no Heckel); В – Taeniocystis mira (Gregarinida)
Правда, метамерия эта является несколько гетерономной, благодаря тому, что камеры не вполне равны между собой, а непрерывно и постепенно увеличиваются от одного конца раковины к другому, так как каждая вновь образующаяся камера несколько больше предыдущей. Впрочем, подобная незначительная степень гетерономности, выражающаяся лишь в постепенном нарастании или уменьшении метамер, в той или иной мере встречается у большинства метамерных животных. У большинства других многокамерных корненожек последовательно образующиеся камеры также располагаются вдоль одной линии, но не прямой, а кривой; чаще всего они располагаются по той или иной спирали. Тем не менее и в этих случаях мы можем говорить о метамерии, если несколько расширить это понятие и включить в него метамерию вдоль криволинейной оси. В этих случаях характер кривой, образующей ось метамерии, становится одним из важнейших морфологических признаков, характеризующих план строения представителей различных видов, родов и семейств корненожек. В аналогичной нодозариям форме метамерия встречается и у некоторых радиолярий, например, у Arthrostrobus из отряда Nasselaria Другой тип метамерии встречается у отдельных представителей грегарин. Среди грегарин Taeniocystis mira имеет дейтомерит, разбитый эктоплазматическими перегородками на целый ряд метамерных отрезков (рис. 25, В).Метамерия выражается здесь в наличии перетяжек и эктоплазматических перегородок. На обоих концах тело грегарины заканчивается неметамерными частями – эпимеритом и задним концом тела; это общее несовершенство всякой метамерии. Неравная величина метамер обуславливает слабо выраженную гетерономность метамерии. В одной из метамер лежит ядро грегарины, которого нет в других метамерах, так как оно единственное; в этом проявляется неполнота метамерии, обусловленная неметамерностью ядерного аппарата. По происхождению своей метамерии Taeniocystis mira резко отличается от метамерных корненожек: здесь она возникает всецело в силу метамерной дифференцировки прежде однородного дейтомерита. Наоборот, во всех известных случаях метамерии у Ciliata мы имеем дело с настоящей стробиляцией, т.е. возникновением метамерного целого путем недоведенного до конца поперечного деления (рис. 25, Г).Деление особи происходит настолько быстро, что не успевает оно дойти до конца, как начинается следующее и возникают целые цепочки особей. Возникшие таким образом цепочки представляют нечто непостоянное, так как они непрерывно распадаются, но тем не менее, поскольку деление тоже идет непрерывно, животное почти все время сохраняет вид цепочки, т.е. сохраняет метамерное строение. Такого типа метамерией обладают главным образом паразитические Astomata (отряд Holotricha): Anoplophrya, Discophrya, Butschliellia и прочие. Из свободноживущих инфузорий аналогичные цепочки образует пресноводная планктическая Sphaerobactrum wurduae, из свободноживущих Flagellata – планктическая динофлагеллята Polykrikos, из паразитических динофлагеллят – Haplozoon lineare (рис. 25, Д). В целом можно сказать, что явления метамерии лишь спорадически возникают в различных группах простейших и большой роли в осложнении их организации не играют (за исключением Foraminifera). В противоположность этому среди низших водорослей (группы, близкой к простейшим) метамерия широко распространена и ведет к возникновению нитчатых форм, столь обычных почти во всех классах водорослей. Здесь метамерия во всех случаях появляется в силу неполного деления. Метамерные нитчатые водоросли имеют вид метамерных рядов клеток (Zygnemaceae, многие Ulotrichales и прочие), иногда разветвленных (например, Cladophora и другие). У кишечнополостных метамерия почти не встречается в строении особей, но довольно широко распространена в строении колоний. Проявлением метамерии упорядочения в строении гидрантов является расположение щупалец в несколько венчиков у некоторых Athecata вместо беспорядочного расположения у других представителей этого подотряда или вместо расположения их в один венчик. Метамерное строение колоний наблюдается весьма часто. Наиболее простой случай представляют стробилы дискомедуз и сходные с ними по способу размножения представители Anthozoa. Название стробилы, данное сначала именно размножающейся поперечным делением личинке дискомедуз, впоследствии стало употребляться в качестве термина для обозначения всякой временной или постоянной колонии, возникающей путем неполного поперечного деления. Наиболее распространенный тип метамерии в колониях кишечнополостных представляют перистые моноподиальные колонии некоторых гидроидов (например, Pennaria, многие Sertulariidae и другие) и Octocorallia. В этих колониях веточки отходят от главного ствола одна против другой и места их отхождения разделены правильными междоузлиями. Здесь можно говорить о настоящей метамерии колонии, и притом метамерии порою очень сложной. Действительно, сами метамерно отходящие веточки, в свою очередь, могут иметь такое же метамерное строение благодаря метамерному расположению сидящих на их гидрантов или ветвлений следующего, высшего порядка. Однако эта сложная метамерия колоний гидроидов представляет сравнительно узкий интерес, и мы на ней подробнее останавливаться не будем. Метамерное строение имеют также и колонии сифонофор-сифонант. Их метамерность бывает выражена при длинном столоне и стирается у форм с укороченным столоном.
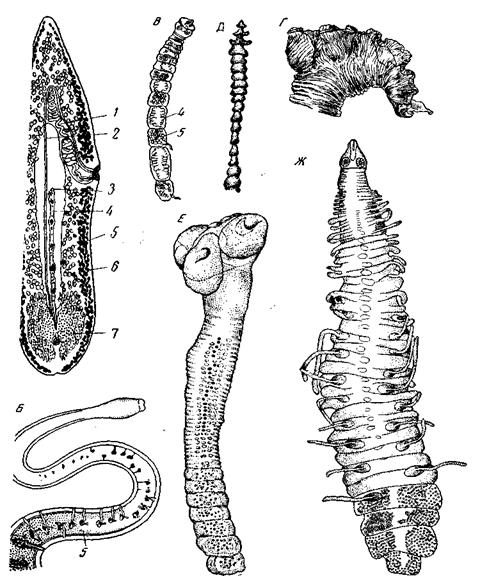
Рис. 26. Формы метамерии турбеллярий А – Microslomum groenlandicum (Macrostomida), цепочка из четырех зооидов (I–IV) (по Graft); Б – Procerodes lobata (Triclada Maricola) (no Lang) 1 – передняя ветвь кишечника у Procerodes, предротовой выступ кишечника у Мicrostomum;
Метамерия сифонант – резко гетерономная благодаря наличию в верхней части столона колоколов и в нижней части – кормидиев. Лучше всего она выражена в нижней части столона (сифоносоме) каликофорид с их упорядоченными кормидиями, разделенными длинными междоузлиями. В отличие от стробил, где метамерия является прямым следствием поперечного деления, в остальных колониях кишечнополостных, возникающих путем почкования, метамерия всегда является метамерией упорядочения. Действительно, почки могут возникать и без всякого определенного порядка и, во всяком случае, без правильной метамерии, что мы и наблюдаем во многих колониях полипов, например, у Alcyonium (Alcyonaria) или Corallium (Gorgonaria), и в этом случае колония не имеет метамерного строения. Гораздо большее значение, чем у особей кишечнополостных, приобретает метамерия у двусторонне-симметричных многоклеточных. Уже у самых примитивных из Bilateria, у плоских червей, мы встречаемся с обеими главными формами метамерии – путем упорядочения и путем стробиляции. Прекрасным примером возникновения метамерии путем упорядочения в расположении органов является морская турбеллярия Procerodes lobata, изученная A. Lang (1881) под названием Gunda segmentata (рис. 26, Б).Lang открыл высокую степень метамерности этого животного и положил ее в основу предлагаемого им объяснения метамерии всех червей вообще. Метамерия Pr. lobata заключается в следующем. Кишечник этого вида, как и у всех триклад, имеет разветвленное строение, причем передняя ветвь дает разветвления направо и налево, а обе задние ветви дают разветвления в сторону наружного края тела. У Pr. lobata все эти веточки имеют совершенно правильное расположение: веточки правой и левой сторон лежат симметрично друг против друга, и все пары лежат на равных расстояниях одна позади другой. Благодаря такому расположению ветвей кишечника половые железы, лежащие в свободных промежутках между ними, в свою очередь, приобретают столь же правильное расположение. Таким образом, метамерными оказываются не только ветви кишечника, но также семенники и желточники. Нервный аппарат состоит из мозга и трех пар продольных стволов; последние на всем своем протяжении соединены между собой комиссурами, кольцевыми и дорсовентральными, которые также располагаются в промежутках между разветвлениями кишечника и в силу этого приобретают такое же метамерное расположение, как и эти последние. Выделительный аппарат Рг. lobata состоит, по Лангу, из двух пар продольных стволов – дорсальной и вентральной. Оба ствола каждой стороны почти в каждом из промежутков между ветвями кишечника соединяются дорсо-вентральной комиссурой. По-видимому, более или менее метамерное расположение имеют и поперечные комиссуры между дорсальными стволами, описанные. От каждого спинного ствола в каждом промежутке отходит по терминальной веточке, которая открывается на спинной стороне 2–4 порами. Таким образом, и выделительный аппарат приобретает метамерное строение. Наконец, и дорсовентральная мускулатура, также всецело приуроченная к промежуткам между выступами кишечника, оказывается вследствие этого метамерной. Итак, метамерия Pr. lobata охватывает почти всю внутреннюю организацию животного. Наоборот, во внешнем виде последнего она не проявляется. Осью метамерии является здесь главная ось тела. Неполнота метамерии Pr. lobata обусловлена присутствием неметамерных органов – глотки и совокупительного аппарата и вызванным присутствием этих органов расщеплением задней половины кишечника на две главные ветви (в то время как передняя половина кишечника остается не расщепленной). Возникновение метамерии Pr. lobata путем метамерного упорядочения органов не вызывает никакого сомнения. У очень многих триклад боковые разветвления кишечника отходят без особой правильности, беспорядочно разбросаны фолликулы желточников и семенники и нервный аппарат также не представляет метамерности. И наряду со случаями такого полного отсутствия метамерии можно подобрать целый ряд форм, у которых все эти органы приобретают все большую правильность расположения и расположение различных аппаратов оказывается все более согласованным. Крайний случай такого метамерного упорядочения почти всex органов тела и представляет Pr. lobata. Менее полная метамерия такого же типа свойственна очень многим трикладам, а из представителей других отрядов турбеллярий – многим Polyclada и Alloeocoela, главным образом лентовидно-вытянутым формам. Rhabdocoela почти лишены метамерии упорядочения благодаря олигомеризации всех своих органов, выражающейся в низведении числа половых желез каждого сорта до одной или двух, уменьшении числа поперечных комиссур нервного аппарата, уменьшении числа отверстий выделительного аппарата до одного или двух и т.д. Еще беднее метамерией ближайшие родичи прямокишечных – сосальщики. Зато в отрядах Macrostomida и Notandropora встречаются случаи бесполого размножения путем поперечного деления, и притом в форме паратомии[1], ведущей к образованию цепочек особей (рис. 26, А).Последние являются зачаточными колониями явно метамерного строения. Таким образом, у этих турбеллярий, как и у Astomata среди инфузорий или у Gonactinia среди Anthozoa, метамерия свойственна не особям, а колониям, и возникает путем не доведенного до конца поперечного деления. Среди Notandropora паратомия свойственна в той или иной мере всем иx представителям, среди Macrostomida – представителям семейства Microstomidae. И хотя оба отряда мало родственны между собой, паратомия встречается у них в довольно сходных формах. У родов Stenostomum Notandropora) и Microstomum (Microstomidae) рост в длину и закладка новых зооидов идут так быстро, что далеко опережают деления, вследствие чего возникают цепочки из 4, 8 и даже 16 особей. При этом растут и делятся все зооиды цепочки в совершенно равной мере. Никакого особого места новообразования зооидов здесь нет. Этим цепочка стеностомид или микростомид существенно отличается от типичной стробилы дискомедуз, у которой новообразование эфир происходит всегда в одном месте, в оральной части первичного зооида – сцифистомы, тогда как обособившиеся эфиры уже больше в длину не растут и не делятся. Итак, в целом мы можем сказать, что в строении и эволюции турбеллярий метамерия играет умеренную роль, а у сосальщиков – почти никакой. Но зато в классе лентецов (Cestodes) метамерия приобретает колоссальное значение. Как известно, Cestodes распадаются на три подкласса Gyrocotyloidea, Cestodaria и Cestoda. Gyrocotyloidea и Cestodaria являются животными неметамерными или не более метамерными, чем Rhabdocoela, с которыми они, вероятно, связаны. Подобно последним, цестодарии обладают единственным набором половых органов. Среди Cestoda на такой ступени стоит только семейство Caryophyllaeidae, прежде относимое к Cestodaria, но затем причисленное к отряду Pseudophyllidea ввиду несомненной близости к последним. Все остальные Cestoda метамерны и большей частью имеют членистое тело, состоящее из сколекса и длинной цепи члеников, или проглоттид. В большинстве случаев проглоттиды хорошо различимы извне благодаря насечкам или перетяжкам, их разделяющим. Каждая проглоттида содержит полный набор половых органов, вполне равноценный всему половому аппарату какой-либо особи Cestodaria. Кроме того, каждая проглоттида содержит поперечные комиссуры нервного и выделительного аппаратов. Между собой проглоттиды различаются только размерами и степенью развития; впереди, вблизи зоны роста, находятся молодые проглоттиды небольших размеров и с неразвитым половым аппаратом; по мере образования новых проглоттид прежде образовавшиеся сдвигаются все дальше кзади, и в то же время они растут и развиваются. Зрелые проглоттиды у большинства цестод отрываются. Итак, метамерия цестод оказывается весьма полной, охватывая все системы органов. Она является, по существу, гомономной (за немногими исключениями, о которых см. ниже), причем гомономность эта нарушается лишь различным возрастом и размерами проглоттид. Неметамерным образованием является только сколекс, и, пожалуй, последняя, старейшая из проглоттид, несущая выделительное отверстие. Судьба зрелых проглоттид у различных цестод различна. 1. Анаполитические стробилы: зрелые проглоттиды не отделяются, большей частью проглоттиды бывают внешне не обособлены. Роды Schistocephalus, Ligula (отряд Pseudophyllidea) и семейство Proteocephalidae (отряд Tetraphyllidea). 2. Аполитические (apo-lytica): стробилы с отделяющимися проглоттидами. Аполитические стробилы, в свою очередь, делятся на: а) гипераполитические, свойственные некоторым Tetraphyllidea, например Т'rilocularia: проглоттиды отделяются очень рано, при едва заложившихся половых органах, и дальше продолжают расти и развиваться, ведя самостоятельный образ жизни в кишечнике хозяина; б) эваполитические, свойственные многим Tetraphyllidea и Tetrarhynchidea: проглоттиды отделяются, имея уже зрелый половой аппарат, но продолжают еще свое развитие в кишечнике хозяина; в) аполитические в тесном смысле слова: отделяются лишь вполне зрелые проглоттиды, набитые яйцами; этот тип свойствен большинству цестод; г) псевдаполитические: проглоттиды проделывают все свое развитие и размножение, не отрываясь от стробилы, отрываются лишь умирающие членики с истощенным половым аппаратом; этот тип стробилы свойствен некоторым Diphyllobothriidae (Pseudophyllidea); он представляет переход к анаполитическим стробилам. Итак, мы встречаемся у цестод с различной степенью обособленности проглоттид. С одной стороны, у анаполитических форм, вроде Ligula или Triaenophorus (Pseudophyllidea) (рис. 27, Б),проглоттиды внешне не обособлены, метамерия проявляется только во внутреннем строении и в этом смысле является менее полной; интеграция стробилы более сильная, индивидуальность проглоттид менее выражена. С другой стороны, у гипераполитических форм, вроде Trilocularia или Phyllobothrivm (рис. 27, А),самостоятельность проглоттид достигает максимума, интеграция стробилы минимальна, проглоттиды отделяются очень рано и большую часть своего развития проделывают, живя самостоятельно. И, наконец, большинство цестод образует всевозможные переходы между этими двумя крайними типами. В связи с таким многообразием типов метамерии цестод взгляды на ее природу также различны. Существуют две теории метамерии цестод. Старые авторы, вплоть до середины XIX в., не сомневались в том, что каждый лентец представляет одно животное, одну особь. Однако в середине прошлого века Однако в конце XIX в. ряд авторов выступил с резкой критикой стробилярной теории, выдвигая на ее место так называемую метамерную теорию строения цестод. Эта теория отрицает наличие чередования поколений у цестод, рассматривает всю цестоду (сколекс и проглоттиды) как одно целое, а проглоттиды рассматривает как членики, возникшие вследствие вторичной метамеризации первоначально единого и нерасчлененного существа. Метамерия цестод возникает, по их мнению, путем умножения половых аппаратов и упорядочения их расположения. В настоящее время эти взгляды являются скорее господствующими, и большинство авторов считает цестоду за особь, а не за колонию. В доказательство своих взглядов сторонники стробилярной теории ссылаются главным образом на гипераполитических и эваполитических тетрафиллид и тетраринхид, сторонники метамерной теории – на анапо-литических протеоцефалид и псевдофиллид. Однако мы считаем, что доводы эти одинаково малоубедительны: существование гипераполитических форм так же мало может само по себе служить доказательством стробилярной теории, как и существование анаполитических форм – доказательством метамерной теории.
Рис. 27. Формы метамерии у ленточных червей А – Phyllobothrium (Tetraphyllidea), отделившийся членик, самостоятельно живущий в кишечнике (из Fuhrmann); Б – передний конец Тriaenophorus nodulosu (Pseudophyllidea), проглоттиды неразграничены, метамерия только в расположении половых аппаратов; В – Gynandrotaenia stambriaria (Cyclophyllidea), сложная метамерия, чередование мужских и женских проглоттид; Г – Fimbriaria fasciolaris (Cyclophyllidea), гетерономная метамерия: следующие за сколексом проглоттиды образуют мощный складчатый псевдосколекс; Д – Idiogenes otidis (Cyclophillidea), гетерономная метамерия, передние проглоттиды образуют псевдосколекс, функционально замещающий отсутствующий у взрослой формы сколекс (из Павловского); Е – Neoshrjabinolepis (Cyclophillidea), сложная метамерия, возникающая благодаря объединению групп рядом лежащих проглоттид в метамеры 2-го порядка (по А.А. Спасскому); Ж – Tatria mathevosiianae (Cyclophyllidea), половые отверстия правильно чередуются справа и слева, так что плоскость симметрии стробилы является плоскостью скользящего отражения (по Окорокову) 1 – семевыносящий проток; 2 – влагалище; 3 – отверстие матки; 4 – матка; 5 – семенники; 6 – желточники; 7 – яичник
Действительно, мы видели, что среди цестод встречаются весьма различные степени интеграции цепочки и независимости проглоттид. Если стать на точку зрения стробилярной теории, существование анаполитических форм, вроде Ligula, особых затруднений не создает: Ligula является с этой точки зрения не более как последним членом в ряду постепенной интеграции стробилы и объединения проглоттид. Точно так же, если стать на точку зрения метамерной теории, существование гипераполитических цестод ничему не мешает: Trilocularia, с ее ранним отделением проглоттид, представляет с этой точки зрения лишь последнее звено в эволюции метамерности, существование которого не может противоречить теории. Очевидно, к этому вопросу надо подходить иначе. Можно ли проводить аналогию между метамерией цестод и метамерией Procerodes?Имеет ли место также и у цестод метамерное упорядочение расположения множества гомотипных органов, первоначально разбросанных без всякого порядка? У Pr. lobata метамерное расположение приобрели различные органы, которые в большом числе, но без метамерной правильности, имеются у других триклад. У цестод речь идет в первую очередь о метамерном повторении целых половых аппаратов. Но большого количества беспорядочно разбросанных половых аппаратов не бывает ни у цестод, ни у других плоских червей, и нет оснований постулировать или изобретать такой план строения. Поэтому для объяснения перехода от одиночных цестод к цепочкам метамерная теория вынуждена прибегать к допущению умножения половых аппаратов, а это допущение ведет к сближению метамерной теории со стробилярной. Действительно, даже с точки зрения стробилярной теории каждая проглоттида не повторяет целиком сколекса: ведь присасывательный аппарат сколекса у нее отсутствует. Не регенерирует он и у самостоятельно живущих члеников тетрафиллид. Даже если стать на стробилярную точку зрения, приходится признать, что в связи с образованием колонии индивидуальность отдельной особи – проглоттиды – у цестод настолько подавлена, что каждая из них имеет не все части тела, которые были бы свойственны одиночной особи, а лишь некоторые из них. Итак, обе спорящие теории причиной возникновения метамерии у цестод признают умножение половых аппаратов. Но сторонники метамерной теории считают это явление за «простое» умножение органов, сторонники стробилярной теории – за подавленное деление. В итоге выбор между обеими теориями зависит от того, какое значение мы придаем умножению органов. Мы считаем, что умножение целых больших отделов тела или целых аппаратов в громадном большинстве случаев приходится толковать как недоведенное до конца бесполое размножение, и, в частности, в отношении цестод нет никакого основания для какой-либо другой точки зрения. Поэтому мы считаем, что стробилярная теория глубже и правильнее отражает природу метамерии цестод, что эти последние действительно являются стробилами. Однако это не решает вопроса, что примитивней, Ligula или Trilocularia; легло ли в основу образования стробил цестод подавленное деление типа умножения органов, с последующим возникновением терминального роста и эмансипацией проглоттид, или же настоящая паратомия, с последующей интеграцией стробилы. Как выше было указано, метамерия цестод, как правило, гомономная. Однако встречаются и случаи гетерономной метамерии. Сюда относятся прежде всего случаи образования прикрепительного органа – псевдосколекса – за счет нескольких передних проглоттид в помощь (рис. 27, Г)или в замену (рис. 27, Д)сколексу. Другой тип гетерономной метамерии представляет Gynandrotaenia stammeri (Cyclophyllidea), представляющая правильное чередование раздельнополых мужских и женских проглоттид (рис. 27, В).Правильность чередования обоих видов проглоттид позволяет рассматривать каждую пару соседних проглоттид как метамеру высшего порядка и говорить о наличии в данном случае гомономной метамерии
Поиск по сайту: |