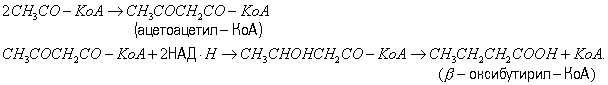|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
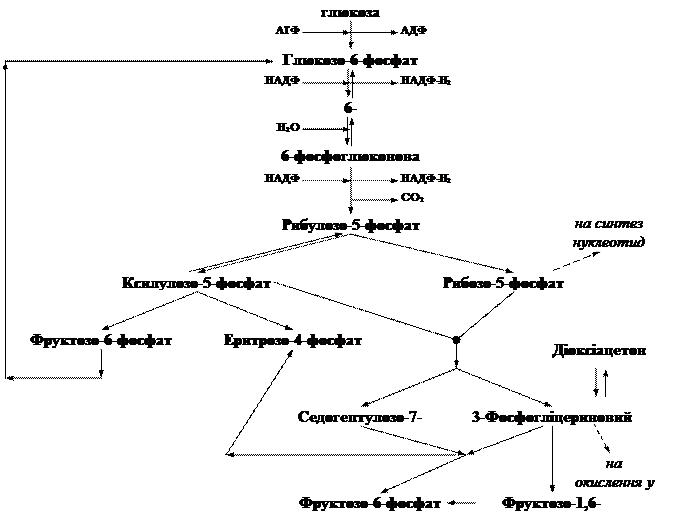
Пентозний цикл (пентозофосфатний або гексозомонофосфатний шлях)
Цей цикл забезпечує організм пентозами, які необхідні для синтезу нуклеїнових кислот тощо. У ссавців активність пентозного циклу відносно висока у печінці, серці, надниркових залозах, ембріональній тканині. Пентозний шлях активно реалізується в еритроцитах людини. Існує група спадкових хвороб людини, при яких активність деяких ферментів цього шляху знижена або взагалі відсутня. У таких хворих спостерігається гемоліз – руйнування еритроцитів з виділенням з них гемоглобіну, що веде до розвитку анемії. Стан різко погіршується під впливом деяких ліків (протималярійних препаратів). В Африці та Азії від цих хвороб страждають мільйони людей. Гексоза, перетворюючись в монофосфат, зразу окислюється в глюконову кислоту. Потім через ряд проміжних стадій утворюються фруктозомонофосфат та фосфогліцериновий альдегід. У подальшому ці продукти піддаються тим же перетворенням, що і при гліколітичному розпаді.
Основне призначення пентозофосфатного циклу: · регенерувати у цитоплазмі відновлювач у формі НАДФ×Н2, що використовується при синтезі жирних кислот і стероїдів; · в ньому синтезуються пентози, особливо D-рибоза, яка необхідна для синтезу нуклеїнових кислот; · участь в утворенні глюкози з СО2 в темновій фазі фотосинтезу.
Важливе значення ферментів пентозофосфатного циклу полягає в тому, що вони разом з ферментами гліколізу створюють умови для взаємоперетворень 3-х, 4-х, 5-ти, 6-ти і 7-вуглецевих сахарів шляхом оборотного перенесення дво- або тривуглецевих фрагментів (глікоальдегідних або діоксіацетонових груп). Отже, глюкозофосфатний шлях (пентозофосфатного циклу) є місцем перехрещення метаболічних шляхів, де утворюється значна кількість різних продуктів обміну речовин. Ферментами фосфоглюконатного шляху може здійснюватися повне окислення глюкозо-6-фосфату до СО2 за таким рівнянням: Глюкозо-6-фосфат + 12 НАДФ ® 6 СО2 + 12НАДФ×Н2 + Рн
Гліоксилатний цикл У тварин він відсутній. Поширений у проростаючому насінні олійних культур. Таким шляхом із ацетильних груп, джерелом яких є жирні кислоти запасних триацилгліцеридів, утворюється глюкоза. Цей цикл є модифікацією циклу лимонної кислоти. Суть його полягає в тому, що ізолимонна кислота під впливом ізоцитрат-ліази розкладається на янтарну та гліоксилеву кислоти. Остання, взаємодіючи з ацетил-КоА під впливом малат-синтази, утворює яблучну кислоту. Яблучна кислота піддається подальшому окисленню, як у циклі Кребса. Жирним виділені реакції, які відрізняють цей цикл від цикла Кребса.
Біосинтез вуглеводів Біосинтез вуглеводів проходить в основному двома шляхами. Перший характерний для автотрофних фотосинтезуючих організмів, які утворюють гексози з вуглекислого газу та води за участі енергії сонячного випромінювання. З цих гексоз далі синтезуються ди- і трисахариди. Другий – для гетеротрофних організмів. Вони такої здатності не мають. Утворення вуглеводів в них відбувається шляхом перетворення органічних сполук, які потрапляють з їжею, а також шляхом синтезу з проміжних сполук, що утворюються в процесі обміну речовин. 1. Глюконеогенез– це процес синтезу глюкози з невуглеводних попередників. Шляхи утворення вуглеводів з органічних речовин сходяться на стадії ПВК. Багато амінокислот перетворюються у вуглеводи через ПВК (глікогенні амінокислоти). Вуглеводи можуть утворюватись із жирів. Жири розщеплюються на гліцерин та жирні кислоти. Останні окисляються у ацетил-КоА. Гліцерин та ацетил-КоА перетворюються у ПВК. Подальший синтез іде традиційним шляхом. Більшість реакцій глюконеогенезу є зворотніми реакціями гліколізу за винятком трьох – гексокіназної, фосфофруктокіназної і піруваткіназної. Ці реакції необоротні, тому в процесі глюконеогенезу на цих стадіях використовуються інші ферменти. Утворення вуглеводів з інших речовин можна представити:
Взаємоперетворення гексоз відбувається через утворення їх фосфорних ефірів. 2. Біосинтез дисахаридів – здійснюється за реакціями трансглікозування. При цьому процес перенесення глікозильного залишку на один моносахариди проходить з фосфорного ефіру другого моносахариду. Реакція каталізується глікозилтрансферазою.
Таким способом синтез відбувається у мікроорганізмів. Також сахароза синтезується шляхом взаємодії уридинфосфосфоглюкози з фруктозою під впливом сахаросинтази. 3. Біосинтез пентоз – пентозофосфатний шлях. 4. Біосинтез крохмалю у рослинах іде у темновій фазі фотосинтезу.
Брожение, процесс анаэробного расщепления органических веществ, преимущественно углеводов, происходящий под влиянием микроорганизмов или выделенных из них ферментов. В ходе Б. в результате сопряженных окислительно-восстановительных реакций освобождается энергия, необходимая для жизнедеятельности микроорганизмов, и образуются химические соединения, которые микроорганизмы используют для биосинтеза аминокислот, белков, органических кислот, жиров и др. компонентов тела. Одновременно накапливаются конечные продукты Б. В зависимости от их характера различают Б. спиртовое, молочнокислое, маслянокислое, пропионовокислое, ацетоно-бутиловое, ацетоно-этиловое и др. виды. Характер Б., его интенсивность, количественные соотношения конечных продуктов, а также направление Б. зависят от особенностей его возбудителя и условий, при которых Б. протекает (pH, аэрация, субстрат и др.). Спиртовое Б. В 1836 французский учёный Каньяр де ла Тур установил, что спиртовое Б. связано с ростом и размножением дрожжей. Химическое уравнение спиртового Б.: C6H12O6 ® 2C2H5OH + 2CO2 было дано французскими химиками А. Лавуазье (1789) и Ж. Гей-Люссаком (1815). Л. Пастер пришёл к выводу (1857), что спиртовое Б. могут вызывать только живые дрожжи в анаэробных условиях («брожение — это жизнь без воздуха»). В противовес этому немецкий учёный Ю. Либих упорно настаивал на том, что Б. происходит вне живой клетки. На возможность бесклеточного спиртового Б. впервые (1871) указала русский врач-биохимик М. М. Манассеина. Немецкий химик Э. Бухнер в 1897, отжав под большим давлением дрожжи, растёртые с кварцевым песком, получил бесклеточный сок, сбраживающий сахар с образованием спирта и CO2. При нагревании до 50°C и выше сок утрачивал бродильные свойства. Всё это указывало на ферментативную природу активного начала, содержащегося в дрожжевом соке. Русский химик Л. А. Иванов обнаружил (1905), что добавленные к дрожжевому соку фосфаты в несколько раз повышают скорость Б. Исследования отечественных биохимиков А. И. Лебедева, С. П. Костычева, Я. О. Парнаса и немецких биохимиков К. Нейберга, Г. Эмбдена, О. Мейергофа и др. подтвердили, что фосфорная кислота участвует в важнейших этапах спиртового Б.
В дальнейшем многие исследователи детально изучили ферментативную природу и механизм спиртового Б. (см. схему). Первая реакция превращения глюкозы при спиртовом Б. — присоединение к глюкозе под влиянием фермента глюкокиназы остатка фосфорной кислоты от аденозинтрифосфорной кислоты (АТФ, см. Аденозинфосфорные кислоты). При этом образуются аденозиндифосфорная кислота (АДФ) и глюкозо-6-фосфорная кислотата. Последняя под действием фермента глюкозофосфати-зомеразы превращается в фруктозо-6-фосфорную кислоту, которая, получая от новой молекулы АТФ (при участии фермента фосфофруктокиназы) ещё один остаток фосфорной кислоты, превращается в фруктозо-1,6-дифосфорную кислоту. (Эта и следующая реакции, обозначенные встречными стрелками, обратимы, т. е. их направление зависит от условий — концентрации фермента, pH и др.) Под влиянием фермента кетозо-1-фосфатальдолазы фруктозо-1,6-дифосфорная кислота расщепляется на глицеринальдегидфосфорную и диоксиацетонфосфорную кислоты которые могут превращаться друг в друга под действием фермента триозофосфатизомеразы. Глицеринальдегидфосфорная кислота, присоединяя молекулу неорганической фосфорной кислоты и окисляясь под действием фермента дегидрогеназы фосфоглицеринальдегида, активной группой которого у дрожжей является никотинамидадениндинуклеотид (НАД), превращается в 1,3-дифосфоглицериновую кислоту. Молекула диоксиацетонфосфорной кислоты под действием триозофосфатизомеразы даёт вторую молекулу глицеринальдегидфосфорной кислоты, также подвергающуюся окислению до 1,3-дифосфоглицериновой кислоты; последняя, отдавая АДФ (под действием фермента фосфоглицераткиназы) один остаток фосфорной кислоты, превращается в З-фосфоглицериновую кислоту, которая под действием фермента фосфоглицеро-мутазы превращается в 2-фосфоглицериновую кислоту, а она под влиянием фермента фосфопируват-гидратазы — в фосфоенол-пировиноградную кислоту. Последняя при участии фермента пируваткиназы передаёт остаток фосфорной кислоты молекуле АДФ, в результате чего образуется молекула АТФ и молекула енолпировиноградной кислоты, которая весьма нестойка и переходит в пировиноградную кислоту. Эта кислота при участии имеющегося в дрожжах фермента пируватдекарбоксилазы расщепляется на уксусный альдегид и двуокись углерода. Уксусный альдегид, реагируя с образовавшейся при окислении глицеринальдегидфосфорной кислоты восстановленной формой никотинамидадениндинуклеотида (НАД-Н), при участии фермента алкогольдегидрогеназы превращается в этиловый спирт. Суммарно уравнение спиртового Б. может быть представлено в следующем виде: C6H12O6 + 2H3PO4 + 2АДФ ® 2CH3CH2OH + 2CO2 + 2АТФ. Т. о., при сбраживании 1 моля глюкозы образуются 2 моля этилового спирта, 2 моля CO2, а также в результате фосфорилирования 2 молей АДФ образуются 2 моля АТФ. Термодинамические расчёты показывают, что при спиртовом Б. превращение 1 моля глюкозы может сопровождаться уменьшением свободной энергии примерно на 210 кдж (50 000 кал), т. е. энергия, аккумулированная в 1 моле этилового спирта, на 210 кдж (50 000 кал) меньше энергии 1 моля глюкозы. При образовании 1 моля АТФ (макроэргических — богатых энергией фосфатных соединений) используется 42 кдж (10 000 кал). Следовательно, значительная часть энергии, освобождающейся при спиртовом Б., запасается в виде АТФ, обеспечивающей разнообразные энергетические потребности дрожжевых клеток. Такое же биологическое значение имеет процесс Б. и у др. микроорганизмов. При полном сгорании 1 моля глюкозы (с образованием CO2 и H2O) изменение свободной энергии достигает 2,87 Мдж (686 000 кал). Иначе говоря, дрожжевая клетка использует лишь 7% энергии глюкозы. Это показывает малую эффективность анаэробных процессов по сравнению с процессами, идущими в присутствии кислорода. При наличии кислорода спиртовое Б. угнетается или прекращается и дрожжи получают энергию для жизнедеятельности в процессе дыхания. Наблюдается тесная связь между Б. и дыханием микроорганизмов, растений и животных. Ферменты, участвующие в спиртовом Б., имеются также в тканях животных и растений. Во многих случаях первые этапы расщепления сахаров, вплоть до образования пировиноградной кислоты, — общие для Б. и дыхания. Большее значение процесс анаэробного распада глюкозы имеет и при сокращении мышц (см. Гликолиз), первые этапы этого процесса также сходны с начальными реакциями спиртового Б. Сбраживание углеводов (глюкозы, ферментативных гидролизатов крахмала, кислотных гидролизатов древесины) используется во многих отраслях промышленности: для получения этилового спирта, глицерина и др. технических и пищевых продуктов. На спиртовом Б. основаны приготовление теста в хлебопекарной промышленности, виноделие и пивоварение. Молочнокислое Б. Молочнокислые бактерии подразделяют на 2 группы — гомоферментативные и гетероферментативные. Гомоферментативные бактерии (например, Lactobacillus delbrückii) расщепляют моносахариды с образованием двух молекул молочной кислоты в соответствии с суммарным уравнением: C6H12O6 = 2CH3CHOH·COOH. Гетероферментативные бактерии (например, Bacterium lactis aerogenes) ведут сбраживание с образованием молочной кислоты, уксусной кислоты, этилового спирта и CO2, а также образуют небольшое количество ароматических. веществ — диацетила, эфиров и т.д. При молочнокислом Б. превращение углеводов, особенно на первых этапах, близко к реакциям спиртового Б., за исключением декарбоксилирования пировиноградной кислоты, которая восстанавливается до молочной кислоты за счёт водорода, получаемого от НАД-Н. Гомоферментативное молочнокислое Б. используется для получения молочной кислоты, при изготовлении различных кислых молочных продуктов, хлеба и в силосовании кормов в сельском хозяйстве. Гетероферментативное молочнокислое Б. происходит при консервировании различных плодов и овощей путём квашения. Маслянокислое Б. Сбраживание углеводов с преимущественным образованием масляной кислоты производят многие анаэробные бактерии, относящиеся к роду Clostridium. Первые этапы расщепления углеводов при маслянокислом Б. аналогичны соответстветственным этапам спиртового Б., вплоть до образования пировиноградной кислоты, из которой при маслянокислом Б. образуется ацетил-кофермент A (CH3CO-KoA). Ацетил-KoA может служить предшественником масляной кислоты, подвергаясь следующим превращениям:
Маслянокислое Б. применялось для получения масляной кислоты из крахмала. Ацетоно-бутиловое Б. бактерии Clostridium acetobutylicum сбраживают углеводы с преим. образованием бутилового спирта (CH3CH2CH2CH2OH) и ацетона (CH3COCH3). При этом образуются также в сравнительно небольших количествах водород, CO2, уксусная, масляная кислоты, этиловый спирт. Первые этапы расщепления углеводов те же, что и при спиртовом Б. Бутиловый спирт образуется путём восстановления масляной кислоты: CH3CH2CH2COOH + 4H = CH3CH2CH2CH2OH + H2O. Ацетон же образуется декарбоксилированием ацетоуксусной кислоты, которая получается в результате конденсации двух молекул уксусной кислоты. Исследованиями В. Н. Шапошникова показано, что ацетоно-бутиловое Б. (как и ряд др., например пропионовокислое, маслянокислое) в опытах с растущей культурой происходит в две фазы. В первую фазу Б. параллельно с нарастанием биомассы накапливаются уксусная и масляная кислоты; во вторую фазу образуются преимущественно ацетон и бутиловый спирт. При ацетоно-бутиловом Б. сбраживаются моносахариды, дисахариды и полисахариды — крахмал, инсулин, но не сбраживаются клетчатка и гемицеллюлоза. Ацетоно-бутиловое Б. использовалось для промышленного получения бутилового спирта и ацетона, применяемых в химической и лакокрасочной промышленности (см. также Ацетоно-бутиловое брожение и Ацетоно-этиловое брожение). Сбраживание белков. Некоторые бактерии из рода Clostridium — гнилостные анаэробы — способны сбраживать не только углеводы, но и аминокислоты. Эти бактерии более приспособлены к использованию белков, расщепляемых ими при помощи протеолитических ферментов до аминокислот, которые затем подвергаются Б. Процесс сбраживания белков имеет значение в круговороте веществ в природе (см. Гниение). Пропионовокислое Б. Основные продукты пропионовокислого Б., вызываемого несколькими видами бактерий из рода Propionibacterium, — пропионовая (CH3CH2OH) и уксусная кислоты и CO2. Химизм пропионовокислого Б. сильно изменяется в зависимости от условий. Это, по-видимому, объясняется способностью пропионовых бактерий перестраивать обмен веществ, например в зависимости от аэрации. При доступе кислорода они ведут окислительный процесс, а в его отсутствии расщепляют гексозы путём Б. Пропионовые бактерии способны фиксировать CO2, при этом из пировиноградной к-ты и CO2 образуется щавелевоуксусная к-та, превращающаяся в янтарную к-ту, из которой декарбоксилированием образуется пропионовая к-та:
Существуют Б., которые сопровождаются и восстановительными процессами. Примером такого «окислительного» Б. служит лимоннокислое Б. Многие плесневые грибы сбраживают сахара с образованием лимонной кислоты. Наиболее активные штаммы Aspergillus niger превращают до 90% потребленного сахара в лимонную кислоту. Значительная часть лимонной кислоты, используемой в пищевой промышленности, производится микробиологическим путём — глубинным и поверхностным культивированием плесневых грибов. Иногда по традиции и чисто окислительные процессы, осуществляемые микроорганизмами, называется Б. Примерами таких процессов могут служить уксуснокислое и глюконовокислое Б. Уксуснокислое Б. Бактерии, относящиеся к роду Acetobacter, окисляют этиловый спирт в уксусную кислоту в соответствии с суммарной реакцией:
Промежуточное соединение при окислении спирта в уксусную кислоту — уксусный альдегид. Многие уксуснокислые бактерии, кроме окисления спирта в уксусную кислоту, осуществляют окисление глюкозы в глюконовую и кетоглюконовую кислоты. Глюконовокислое Б. осуществляют и некоторые плесневые грибы, способные окислять альдегидную группу глюкозы, превращая последнюю в глюконовую кислоту:
Кальциевая соль глюконовой кислоты служит хорошим источником кальция для людей и животных.
Лит.: Шапошников В. Н., Техническая микробиология, М., 1948; Прескот С., Дан С., Техническая микробиология, пер. с англ., М., 1952; Пастер Л., Избр. труды, пер. с франц., т. 1—2, М., 1960; Кретович В. Л., Основы биохимии растений, 4 изд., М., 1964; Фробишер М., Основы микробиологии, пер. с англ., М., 1965; Фердман Д. Л., биохимия, М., 1966; Работнова И. Л., Общая микробиология, М., 1966.
Поиск по сайту: |



 Так, при синтезі сахарози сахарозоглюкозилтрансфераза каталізує перенесення залишку глюкози з глюкозо-1-фосфату на молекулу фруктози:
Так, при синтезі сахарози сахарозоглюкозилтрансфераза каталізує перенесення залишку глюкози з глюкозо-1-фосфату на молекулу фруктози: