|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Особенности развития головы и головного мозга
Голова человека, как и голова всех позвоночных, представляет собой обособленный передний (у человека — верхний) отдел тела, своеобразно измененный в соответствии с теми функциональными задачами, которые он выполняет. В процессе развития головы происходит формирование головного мозга и органов чувств, а также скелета головы — черепа, состоящего из мозгового и висцерального отделов. Мозговой череп окружает головной мозг и связанные с ним органы чувств, обеспечивая их защиту и опору (рис. 52). Висцеральный череп развивается в связи с формированием челюстного аппа-
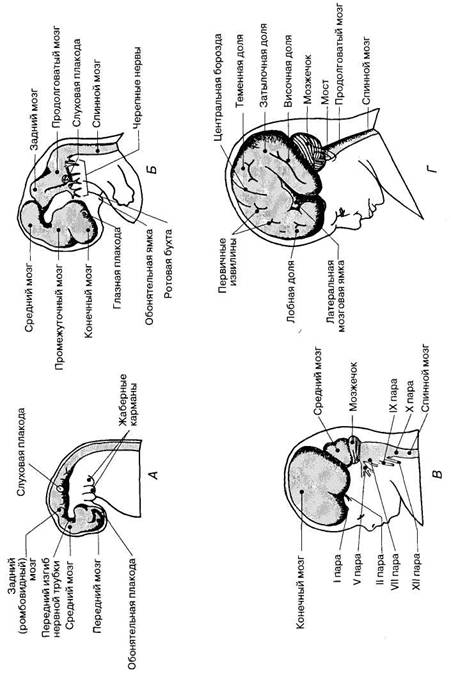
Развитие головы связано с утолщением переднего конца нервной трубки эмбриона, формирующейся из эктодермальной нервной пластинки в результате слияния ее боковых валиков (нейруляции). На 4-й неделе эмбрионального развития из переднего (рострального) отдела нервной трубки образуются три мозговых пузыря (рис. 53). Первым появляется задний мозговой пузырь — та часть нервной трубки, из которой в дальнейшем образуется задний, или ромбовидный, мозг (rhomben-cephalon), непосредственно соединенный со спинным мозгом. Далее формируется средний мозговой пузырь — зачаток среднего мозга (mesencephalon); наконец, возникает передний мозговой пузырь, или передний мозг (prosencephalon). К концу 4-й недели стенка нервной трубки состоит из трех слоев: краевого (наружного), мантийного (среднего) и эпендимного (внутреннего). Нарастание клеточной массы в нервной трубке происходит за счет внутреннего эпендимного слоя, содержащего нейробласты и спонгиобласты — клетки, способные к размножению. Размножение (пролиферация) и созревание вновь образованных нейробластов и глиоцитов происходит в мантийном слое, а в тонком краевом слое располагаются волокна и верхушечные дендриты созревающих нейронов. Одновременно из клеток эмбриональной мезенхимы, выселяющихся из склеротомов трех-четырех головных сомитов, которые закладываются на уровне заднего мозгового пузыря, начинает формироваться хрящевая капсула. Из нее впоследствии образуются кости основания мозгового черепа. Она состоит из двух хрящевых прехордальных пластинок, расположенных в области переднего мозгового пузыря, и двух парахордальных пластинок, расположенных по бокам от хорды (рис. 54). При срастании этих пластинок между
Рис. 54. Формирование скелета головы.
В области головы не встречаются типичные сомиты (метамеры), подобные туловищным сегментам. Как уже отмечалось, для спинного мозга характерно выраженное сегментарное строение, которое отражает осевую метамерию тела, связанную главным образом с построением двигательного аппарата: сегментарным строением скелета туловища (позвоночного столба и ребер), мышц и спинномозговых нервов. В области головы разделение скелета на отдельные костные сегменты, подобные позвонкам и ребрам, отсутствует; головной мозг также формируется в основном как надсегментарное образование. Исключение составляет задний мозг, которому присущи некоторые черты метамерной организации вследствие его связей с формированием жаберного аппарата и с выходом большей части черепных нервов. Нервные узлы черепных нервов, так же, как и сенсорные узлы спинномозговых нервов, образуются из нервного гребня — длинного тяжа, расположенного по обеим сторонам крыльных пластинок нервной трубки и состоящего из клеток нейроэктодермы. Нервный гребень, доходящий в краниальном отделе нервной трубки до среднего мозга, дает начало периферическим постганглионарным нейронам ресничного, крылонебного, подчелюстного и подъязычного вегетативных узлов и их нервным сплетениям, а также сенсорным нейронам в составе узлов сенсорного (VIII пара) и бранхиогенных (V, VII, IX, X пары) черепных нервов. Сенсорные ганглии черепных нервов связаны также с эктодермальными плакодами, участвующими в формировании органов чувств (органов зрения, слуха, обоняния и вкуса). Из клеток плакод развиваются эпителиальные выстилки наружного слухового хода и наружной поверхности барабанной перепонки, глаза, перепончатого лабиринта и кортиева органа, носослезного канала и околоносовых пазух, слизистой оболочки ротовой и носовой полостей. Из них же формируются эмаль зубов, хрусталик глаза, аденогипофиз, железистая ткань слюнных и слезных желез, вкусовые рецепторы. Соматомоторные (III, IV, VI, XII пары) черепные нервы, а также двигательные волокна в составе бранхиогенных черепных нервов (V, VII, IX, X, XI пары), иннервирующих мышцы жаберного происхождения, формируются аксонами нейронов, развивающихся из клеток базальной пластинки среднего и заднего мозговых пузырей (рис. 55). Парасимпатические преганглионар-ные нейроны вегетативных ядер III, VII, IX и X пар черепных нервов разви-
Рис. 55. Закладка ядер черепных нервов и их локализация в нервной трубке в области ствола мозга. ваются из межуточной зоны, расположенной между крыльной и базальной частями стенок среднего и заднего мозговых пузырей. К концу 3-й недели эмбриогенеза, еще до завершения нейруляции и формирования мозговых пузырей в результате быстрого роста краниального отдела нервной трубки происходит ее изгиб вперед и вверх. Затем формируются и другие изгибы нервной трубки. На 5-й неделе эмбриогенеза число мозговых пузырей увеличивается до пяти (табл. 3). Вначале из заднего мозгового пузыря обособляется задний мозг (metencephalon) и продолговатый мозг (myelencephalon). Чуть позднее из переднего мозгового пузыря формируется конечный мозг (telencephalon) и промежуточный мозг (diencephalon). Конечный мозг сразу же закладывается как парное образование, состоящее из правого и левого полушарий. Последующее развитие мозговых пузырей имеет свои особенности. В начале 5-й недели продолговатый мозг почти не отличается от спинномозгового отдела нервной трубки: он имеет крыльную и базальную пластинки. В толще базальной и крыльной пластинок происходит закладка ядер V—XII пар черепных нервов, причем моторные ядра формируются из материала базальной пластинки, поэтому располагаются медиальнее по отношению к сенсорным ядрам — производным крыльной пластинки. Формирующиеся вегетативные ядра черепных нервов занимают промежу-
В развитии заднего мозга прослеживаются те же изменения в расположении пластинок нервной трубки, что и в продолговатом мозге. Вследствие расхождения крыльных пластинок в стороны здесь также происходит истончение крыши нервной трубки. Миграция клеток крыльной пластинки в область крыши заднего мозга приводит к формированию на 12—16-й неделях червя и полушарий мозжечка. Под мозжечком просматривается полость будущего IV желудочка, дно которой образует ромбовидная ямка. Средний мозг обособляется относительно рано, на 4-й неделе развития. Клетки крыльной пластинки в области среднего мозга мигрируют в дорсальном и- вентральном направлениях. На дорсальной поверхности они дают начало ядрам верхних и нижних холмиков пластинки будущего четверохолмия. Клетки, смещающиеся вентрально, формируют красные ядра и ретикулярные ядра среднего мозга. Из базальной пластинки образуется также ядро
Промежуточный мозг обособляется от переднего мозгового пузыря на 5-й неделе. В ростральной (передней) части нервной трубки, дающей начало конечному и промежуточному мозгу, идет интенсивное увеличение числа нервных клеток. Здесь отсутствует деление на крыльную и базальную пластинки, не происходит формирования черепных нервов, а вся масса нервных клеток идет на развитие многочисленных ядер и коры головного мозга. В промежуточном мозге на 5-й неделе наблюдается начало интенсивного роста мантийного (пролиферативного) слоя, что приводит к значительному утолщению его латеральных стенок; здесь образуются закладки правого и левого таламусов. Еще до обособления промежуточного мозга в задней части переднего мозгового пузыря на его вентролатеральной поверхности появляются парные глазные пузыри, которые растут в сторону эктодермы в форме бокалов. Из стенки глазного бокала сформируются светочувствительные и нервные элементы сетчатки глаза, а из эктодермы в области глазных плакод и окружающей мезенхимы — все остальные структуры глаза. В силу этого зрительный нерв (II пара) не является типичным черепным нервом, а представляет собой специальное образование, связывающее сетчатку с головным мозгом, а сама сетчатка фактически является частью стенки нервной трубки, отделившейся от нее в процессе развития и выселившейся на периферию. Одновременно с развитием конечного мозга начинают формироваться их ростральные выросты, из которых впоследствии образуются обонятельные луковицы и обонятельные тракты (I пара черепных нервов). Они также не являются типичными черепными нервами, а представляют собой проводниковый отдел обонятельного анализатора. Конечный мозг с самого начала развития закладывается как парное образование в виде двух будущих полушарий. Внутри полушарий образуются парные полости — боковые желудочки мозга. В период с 6-й до 12-й недели эмбриогенеза полушария головного мозга интенсивно растут, значительно опережая в росте все другие отделы головного мозга. В результате полушария, как плащ, покрывают эти другие отделы. Часть нейробластов мигрирует к поверхности стенки переднего мозгового пузыря, образуя в дальнейшем кору мозга. К началу 4-го месяца внутриутробной жизни (т. е. чуть более, чем через 100 дней с момента начала развития организма) интенсивное деление нейробластов внутреннего плаща приводит к тому, что у зародыша человека число нервных клеток уже приближается к их числу в коре у новорожденного. Слой, в котором происходит размножение нейробластов, располагается наиболее глубоко, поэтому в процессе развития вновь образованные ней-робласты мигрируют наружу в мантийный слой, где они постепенно приобретают типичную для себя форму и занимают определенное положение в формирующейся коре мозга (рис. 56). Миграцию нейробластов направляют клетки радиальной глии, образующиеся из спонгиобластов эпендимы. Нейроны в процессе миграции как бы «скользят» вдоль отростков этих глиаль-ных клеток из глубины мантийного слоя по направлению к поверхности мозгового пузыря. Нейробласты, мигрировавшие первыми, заполняют сначала
Рис. 56. Миграция нейробластов в процессе формирования коры больших полушарий. глубинные слои будущей коры, а клетки, образовавшиеся позднее, располагаются в более верхних слоях. Таким образом, кора головного мозга формируется как бы «изнутри кнаружи». При этом время образования нейрона определяет его принадлежность к определенным слоям коры, а значит и его функции. Созревание нейробластов происходит гораздо медленнее, чем их пролиферация (накопление количества), и продолжается после рождения в течение первых лет жизни. В коре мозга раньше всего созревают крупные пирамидные клетки ее нижнего этажа (проекционные нейроны), связывающие кору с нижележащими отделами головного и спинного мозга. Их можно наблюдать уже на 8-й неделе эмбриогенеза. К 25-й неделе внутриутробного развития в корковой пластинке плаща можно различить цитоархитектонические слои. Рост и развитие плаща приводит к образованию борозд и извилин (складок) на поверхности конечного мозга, которые формируются на 28—ЗО-й неделях. Окончательное завершение формирования рельефа полушарий наблюдается в постнатальном периоде примерно к 7-8 годам жизни ребенка. Последовательность и степень развития рельефа полушарий (борозд и извилин) программируется генетически, однако его строение в значительной степени зависит также от внешних влияний окружающей среды (соотношения благоприятных и неблагоприятных факторов существования, степени защищенности от влияния неблагоприятных условий, степени вынужденной активности индивидуума, информационной обогащенности среды и т. д.) и от состояния внутренней среды организма (уровня энергетического обмена, соотношения и уровня гормонов в крови и т. д.). Это обусловливает существенные индивидуальные различия в величине площади коры в разных долях и извилинах мозга.
Ствол мозга (truncus encephali) объединяет три отдела головного мозга: продолговатый мозг, мост и средний мозг. Как и для спинного мозга, от которого отходят спинномозговые нервы, для ствола характерно отхождение черепных нервов (с III по XII пару), иннервирующих мускулатуру и кожу головы. внутренние органы и производные жаберного аппарата (рис. 57). Наряду с этим ствол мозга служит тем образованием, через которое осуществляется связь головного мозга со спинным посредством восходящих и нисходящих проводящих путей. Наконец, в стволе мозга находятся также центры, имеющие общеорганизменное значение и связанные с регуляцией дыхания (дыхательный центр), кровообращения (сердечно-сосудистый центр), мышечного тонуса и другие.
Для всех отделов мозга, входящих в состав его ствола, характерно сходное распределение серого и белого вещества. На всем протяжении в нем выделяют крышу (tectum) — скопление серого вещества, расположенное над полостями мозга, покрышку (tegmentum), в которой локализуются ядра черепных нервов, а также проходит ряд проводящих путей восходящего и нисходящего направления, и основание (basis), где сосредоточены нисходящие проводящие пути.
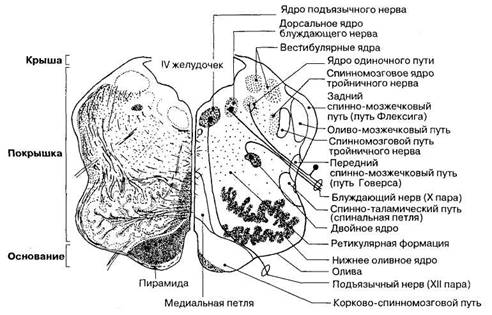
Продолговатый мозг (bulbus, medulla oblongata) представляет собой непосредственное продолжение спинного мозга, поэтому в его строении в большей мере, чем в других отделах ствола мозга, проявляются некоторые черты сходства со спинным мозгом. Верхний расширенный конец продолговатого мозга граничит с нижним краем моста, а нижний соответствует месту выхода корешков I пары шейных спинномозговых нервов. На передней (нижней) поверхности продолговатого мозга проходит передняя срединная щель, являющаяся продолжением аналогичной щели в спинном мозге. По бокам от нее расположены два продольных возвышения — пирамиды. Они состоят из белого вещества и образованы волокнами пирамидных проводящих путей. Часть пирамидных волокон в глубине передней срединной щели переходит на противоположную сторону, образуя перекрест пирамид. Далее волокна из пирамид продолжаются в передние и боковые канатики спинного мозга. Снаружи от пирамид справа и слева находятся возвышения — оливы, внутри каждой из которых заметно скопление серого вещества, образующего нижнее оливное ядро. Они функционально связаны с регуляцией равновесия и работой вестибулярного аппарата. Между пирамидой и оливой расположена передняя латеральная борозда — место выхода корешков подъязычного нерва (XII пара). По дорсальной поверхности продолговатого мозга проходит задняя срединная борозда, являющаяся продолжением одноименной борозды спинного мозга. По бокам от нее идут задние латеральные борозды. Между задней срединной и латеральной бороздами с каждой стороны продолговатого мозга расположены по два утолщения — тонкий и клиновидный бугорки, внутри которых находятся одноименные ядра. На нервных клетках этих ядер заканчиваются волокна тонкого и клиновидного пучков, продолжающихся из спинного в продолговатый мозг. Под задними канатиками внутри нижней трети продолговатого мозга находится центральный канал, являющийся продолжением аналогичного канала спинного мозга. Этот канал открывается в IV желудочек — полость в области ствола мозга. Дно IV желудочка образует ямка ромбовидной формы — ромбовидная ямка. Участки продолговатого мозга, ограниченные латеральными бороздами, — это боковые канатики, которые также являются продолжением боковых канатиков спинного мозга. Волокна из боковых канатиков без резкой границы переходят в нижние ножки мозжечка. Из толщи боковых канатиков выходят корешки языкоглоточного (IX пара), блуждающего (X пара) и добавочного (XI пара) нервов. Внутреннее строение. Для внутреннего строения продолговатого мозга характерно особое распределение серого и белого вещества в крыше, покрышке и основании этого отдела мозга. Крыша продолговатого мозга развита слабо; она лежит выше центрального канала и прикрывает сверху нижнюю часть IV желудочка. Крыша образована тонким слоем клеток, составляющим нижний мозговой парус, — нейро-
Рис. 58. Топография серого и белого вещества в продолговатом мозге: слева показаны пучки нервных волокон, справа — локализация ядер. эпителиальную пластинку, натянутую между ножками клочков мозжечка и представляющую собой остаток стенки первичного мозгового пузыря. Покрышка — наиболее объемная часть продолговатого мозга; она непосредственно образует дно Г/ желудочка (рис. 58). В ее толще расположены нижние оливные ядра и ядра IX, X, XI и XII пар черепных нервов. Эти ядра проецируются на поверхность ромбовидной ямки. В связи с особенностями развития нервной трубки и топографического положения крыльной и базальной пластинок двигательные ядра черепных нервов расположены наиболее медиально, чувствительные — латерально, а вегетативные занимают промежуточное положение. В покрышке продолговатого мозга лежат жизненно важные центры общеорганизменного значения, регулирующие деятельность сердца и кровеносных сосудов (сосудодвигательный центр), акт дыхания (дыхательный центр). Эти центры расположены в вегетативном заднем (дорсальном) ядре блуждающего нерва (X пара). При его повреждении немедленно прекращаются дыхательные движения и регуляция работы сердца, резко падает кровяное давление, что приводит к быстрой смерти. В заднем ядре блуждающего нерва находится также центр рвоты, а в двигательных ядрах IX, X и XII пар черепных нервов — центры чихания и кашля. В покрышке лежат также некоторые ядра ретикулярной формации. Особенности структурной организации продолговатого мозга представлены на рис. 59. В покрышке продолговатого мозга расположен ряд восходящих и некоторых экстрапирамидных нисходящих проводящих путей. В области тонкого и
Рис. 59. Основные компоненты анатомического строения продолговатого мозга. Т — восходящие проводящие пути; 4- — нисходящие проводящие пути. клиновидного бугорков располагаются одноименные ядра, в которых происходит переключение афферентных волокон, несущих проприоцептивные импульсы от всех частей тела, кроме головы. От этих ядер слева и справа отходят внутренние дугообразные волокна, образующие перекрест медиальных петель. Перейдя на противоположную сторону, эти волокна формируют в
Рис. 60. Формирование в продолговатом мозге восходящих нервных путей общей чувствительности — медиальной петли.
В составе нисходящих путей через покрышку продолговатого мозга проходят волокна красноядерно-спинномозгового, тектоспинального, ретику-лоспинального и других нисходящих трактов, направляющиеся в передние и боковые канатики спинного мозга. Ближе к дорсальной поверхности в покрышке проходит парный медиальный продольный пучок волокон, связывающий между собой двигательные ядра шейного отдела спинного мозга, вестибулярные ядра и ядра черепных нервов, управляющих движениями глаз (III, IV, VI пары). Этот пучок обеспечивает согласованные движения глаз при любом пространственном положении головы, а также координирует повороты глаз при движениях головы и шеи в ту или другую сторону. Основание продолговатого мозга представлено пирамидами. Серое вещество в нем отсутствует; здесь проходят нисходящие двигательные корково-спинномозговые пути, направляющиеся в передние и боковые канатики спинного мозга и обеспечивающие сознательное управление движениями. Мост Мост (pons) имеет снизу и спереди вид толстого белого валика, расположенного в средней части ствола мозга. Вверху он граничит с ножками мозга, а внизу — с продолговатым мозгом. На границе между мостом и продолговатым мозгом находятся места выхода VI, VII и VIII пар черепных нервов (см. рис. 57). Значительная часть массы моста состоит из поперечно расположенных, перекрещивающихся между собой нервных волокон, которые идут в обе стороны и направляются к мозжечку, образуя его средние ножки. В месте перехода моста в среднюю ножку мозжечка выходит тройничный нерв (V пара). Задняя (дорсальная) поверхность моста образует верхнюю часть ромбовидной ямки. В ромбовидной ямке на границе моста с продолговатым мозгом хорошо различимы поперечные мозговые полоски, в которых проходят волокна слухового пути. На вентральной поверхности посередине хорошо видна базилярная борозда, в которой лежит базилярная артерия, снабжающая кровью головной мозг. Внутреннее строение моста характеризуется определенной локализацией серого и белого вещества в пределах его крыши, покрышки и основания (рис. 61). Крыша моста так же, как и в продолговатом мозге, развита слабо и образована тонкой нейроэпителиальной пластинкой — верхним мозговым парусом. Он натянут между верхними ножками мозжечка, ограничивающими верхний угол ромбовидной ямки.
Рис. 61. Топография серого и белого вещества на горизонтальном срезе моста: слева показаны пучки нервных волокон, справа — локализация ядер.
Рис. 62. Основные компоненты анатомического строения моста.
Покрышка моста занимает пространство от поверхности ромбовидной ямки до нижней границы трапециевидного тела, образованного поперечными пучками слуховых волокон, располагающихся в толще моста (рис. 62). В покрышке находятся ядра V, VI, VII и VIII пар черепных нервов, проекция которых на поверхность ромбовидной ямки показана на рис. 63. Как и в продолговатом мозге, двигательные ядра черепных нервов расположены медиальнее, а чувствительные — латеральнее остальных ядер. В покрышке моста локализуется парное верхнее оливное ядро, в котором переключается часть слуховых волокон, а также располагаются ядра ретикулярной формации.
Слуховые волокна нейронов дорсального и вентрального улитковых ядер преддверно-улиткового нерва (VIII пара) проходят в поперечном направлении на противоположную сторону моста в составе трапециевидного тела и частично — по дну ромбовидной ямки в мозговых полосках. После перекреста слуховые волокна дугообразно изгибаются и следуют в восходящем направлении к среднему и промежуточному мозгу. Этот изгиб слуховых волокон получил название латеральной петли, так как он лежит снаружи от медиальной петли. Основание моста в отличие от аналогичного образования продолговатого мозга состоит как из белого, так и из серого вещества. Серое вещество представлено собственными ядрами моста. Эти ядра играют роль промежуточных центров связи коры больших полушарий с мозжечком. Большую часть волокон основания моста составляют поперечные волокна, берущие начало на нейронах ядер моста и направляющиеся к мозжечку через его средние ножки. При этом в области моста происходит их перекрест, так что каждое полушарие большого мозга непосредственно влияет на работу контрлатеральной (противоположной) половины мозжечка. В основании моста идут нисходящие эфферентные волокна корково-спинномозговых путей (пирамидных путей). Средний мозг По сравнению с другими отделами ствола мозга средний мозг (mesencephalon) у млекопитающих и человека имеет небольшие размеры. Переднюю (нижнюю) часть его составляют ножки мозга, а заднюю (верхнюю) — пластинка четверохолмия (см. рис. 57). Полостью среднего мозга является узкий канал — водопровод мозга. Внутреннее строение. Для среднего мозга характерно особое распределение серого и белого вещества в крыше, покрышке и основании, образованном ножками мозга (рис. 64).
Рис. 64. Топография серого и белого вещества в среднем мозге: слева показаны пучки нервных волокон, справа — локализация ядер. Крыша среднего мозга, или пластинка четверохолмия, состоит из небольших возвышений — двух верхнихи двух нижних холмиков. Внутри этих холмиков находятся скопления серого вещества — ядра, играющие важную роль в осуществлении зрительных и слуховых рефлексов. В области верхних холмиков лежат подкорковые зрительные центры, в области нижних — подкорковые слуховые центры. Каждый холмик при помощи так называемых ручек связан с коленчатыми телами — образованиями промежуточного мозга. Основание среднего мозга составляют ножки мозга — два толстых белых тяжа, идущих от моста кверху и наружу и затем погружающихся в вещество большого мозга. Между внутренними поверхностями ножек на основании мозга находится лшжножковая ямка и заднее продырявленное вещество. Здесь на вентральной поверхности с внутренней стороны каждой ножки мозга выходит глазодвигательный нерв (III пара). На дорсальной поверхности ствола позади пластинки крыши находится место выхода блокового нерва (IV пара). Посредством черного вещества основание ножки мозга отделено от покрышки. В покрышке среднего мозга под водопроводом мозга в окружающем его сером веществе расположены ядра глазодвигательного и блокового нервов, а также среднемозговое (мезэнцефалическое) ядро тройничного нерва,
Рис. 65. Основные компоненты анатомического строения среднего мозга. продолжающееся сюда из моста. Кроме них серое вещество покрышки образует парное красное ядро (nucl. ruber), которое представляет собой крупное скопление нервных клеток, играющее наряду с другими образованиями ключевую роль в экстрапирамидной системе мозга. Здесь же расположено черное вещество — парное подкорковое ядро, также относящееся к экстрапирамидной системе. В состав серого вещества входят ядра ретикулярной формации. Белое вещество покрышки среднего мозга разнообразно по составу волокон. Здесь проходят восходящие чувствительные и нисходящие двигательные пути (рис. 65). Восходящие сенсорные пути — медиальная, спинальная и тригеминальная петли (проводящие импульсы проприоцептивной и экс-тероцептивной чувствительности) и латеральная (слуховая) петля — идут в толще покрышки ближе к ее латеральной поверхности. Нисходящие пути — тектоспинальный и красноядерно-спинномозговой — занимают в покрышке медиальное положение, так как они образуют перекрест по срединной линии. Тектоспинальный путь1 берет начало от ядер верхних и нижних холмиков в пластинке крыши. Аксоны нейронов, расположенных в этих ядрах, огибают снизу водопровод мозга и переходят на противоположную сторону, образуя дорсальный перекрест покрышки. После перекреста волокна направляются в
В покрышке среднего мозга, как и в других отделах ствола мозга, расположены ядерные и волокнистые структуры ретикулярной формации. В основании ножек мозга проходят нисходящие корково-мосто-мозжечко-вые и пирамидные (корково-спинномозговые и корково-ядерный) пути, связывающие кору головного мозга с мозжечком и спинным мозгом. Ретикулярная формация Ретикулярная формация (formatio reticularis) представляет собой филогенетически более старую и относительно просто организованную нервную сеть с множеством ядерных центров. Ей отводится важная роль в поддержании бодрствующего состояния мозга, а также в механизмах формирования сложно-координированных двигательных актов (таких, как чихание, рвота и т. п.), обеспечивающих защиту организма от воздействий внешней среды, угрожающих его жизнедеятельности. Она работает в функциональном единстве с анализаторными системами и оказывает тонические влияния на ниже- и вышележащие отделы центральной нервной системы. К числу особенностей строения ретикулярной формации следует отнести: • глубокое и диффузное расположение ее ядер в стволе мозга; при про • способность проводить возбуждения в различных, как восходящих, • обслуживание как соматических, так и висцеральных функций; • разнообразие групп ядер по хемоархитектонике (наличие клеточных Ретикулярные ядра группируются в стволе мозга в три билатерально симметричных продольных столба: срединный (ядра шва), медиальный, содержащий ядра с крупными ретикулярными нейронами, и латеральный, содержащий ядра из мелких и средних по размерам нейронов (рис. 66). Множественность и разнообразие групп ядер, входящих в состав ретикулярной формации, позволяют характеризовать ее как неспецифическую по-лисинаптическую систему. В частности, влияние ретикулярной формации на кору больших полушарий осуществляется посредством ретикуло-таламиче-ских путей через группу медиальных ядер таламуса, составляющих так называемую неспецифическую проекционную таламо-кортикальную систему. Волокна ретикуло-таламо-кортикальных путей заканчиваются во всех областях коры (так называемые неспецифические афферентные волокна коры боль-
Рис. 66. Ядра ретикулярной формации в стволе мозга. шого мозга), но наибольшее их число обнаруживают в коре лобной доли. Они проводят грубую (протопатическую) чувствительность от кожного покрова и всех органов, оказывая неспецифическое активирующее воздействие на кору больших полушарий. Их называют неспецифическими афферентными волокнами в отличие от специфических, проводящих более тонкую (эпикрити-ческую) чувствительность к соматосенсорной коре, которая позволяет тонко дифференцировать локализацию и нюансы ощущений. Специфические афферентные волокна заканчиваются на вставочных нейронах IV слоя коры аксосоматическими контактами, обеспечивающими быстрые и дифференцированные ответы нейронов. Неспецифические афферентные волокна в составе восходящей активирующей системы образуют концевые ветвления во всех слоях коры мозга и формируют синаптические контакты на дендри-тах нейронов, что позволяет только модулировать возбудимость нейронов и влиять на их способность к проведению импульсов. Активность как восходящих, так и нисходящих систем поддерживается непрерывным притоком афферентных импульсов, поступающих в ретикулярную формацию по коллатеральным волокнам от сенсорных проводящих путей. Важную роль в поддержании активности ретикулярных ядер играют гуморальные раздражители, по отношению к которым ретикулярная формация обладает высокой чувствительностью, что обеспечивает ее участие в регуляции ряда вегетативных функций. Все двигательные ядра черепных нервов получают от ретикулярных ядер волокна, идущие в составе восходящих и двух нисходящих ретикулярных путей: мосто-спинномозгового {латеральный ретикулоспиналъный путь) и бульбо-спинномозгового {медиальный ретикулоспиналъный путь). Волокна от ретикулярных ядер заканчиваются билатерально на мотоней-
Поиск по сайту: |

 рата, служащего для захвата и первичной механической обработки поступающей в организм пищи; кроме того, висцеральный череп принимает участие в формировании начальных отделов дыхательных путей.
рата, служащего для захвата и первичной механической обработки поступающей в организм пищи; кроме того, висцеральный череп принимает участие в формировании начальных отделов дыхательных путей.
 ними остается отверстие напротив задней части переднего мозгового пузыря — карман Ратке, на месте которого в дальнейшем будет сформировано костное углубление (турецкое седло) для гипофиза. Ростральнее прехордаль-ных пластинок из отдельных мезенхимных зачатков образуются хрящевые носовые капсулы, впоследствии — кости стенок носовой полости. Латераль-нее парахордальньгх пластинок закладываются слуховые капсулы (из них разовьется костный лабиринт внутреннего уха), а вентральнее — хрящи пяти пар висцеральных дуг (бранхиогенный скелет, связанный с формированием жаберного аппарата). Из материала I и II висцеральных дуг и окружающей их мезенхимы будут сформированы кости лицевого черепа: верхняя челюсть, небная и скуловая кости, нижняя челюсть, подъязычная кость и слуховые кости. Хрящевые зачатки последующих висцеральных, или жаберных, дуг послужат для формирования хрящей гортани.
ними остается отверстие напротив задней части переднего мозгового пузыря — карман Ратке, на месте которого в дальнейшем будет сформировано костное углубление (турецкое седло) для гипофиза. Ростральнее прехордаль-ных пластинок из отдельных мезенхимных зачатков образуются хрящевые носовые капсулы, впоследствии — кости стенок носовой полости. Латераль-нее парахордальньгх пластинок закладываются слуховые капсулы (из них разовьется костный лабиринт внутреннего уха), а вентральнее — хрящи пяти пар висцеральных дуг (бранхиогенный скелет, связанный с формированием жаберного аппарата). Из материала I и II висцеральных дуг и окружающей их мезенхимы будут сформированы кости лицевого черепа: верхняя челюсть, небная и скуловая кости, нижняя челюсть, подъязычная кость и слуховые кости. Хрящевые зачатки последующих висцеральных, или жаберных, дуг послужат для формирования хрящей гортани.
 Таблица 3.Развитие отделов головного мозга
Таблица 3.Развитие отделов головного мозга точное положение между моторными и сенсорными. Одновременно с закладкой ядер, а также дифференцировкой клеток нервного гребня происходит развитие самих черепных нервов.
точное положение между моторными и сенсорными. Одновременно с закладкой ядер, а также дифференцировкой клеток нервного гребня происходит развитие самих черепных нервов. глазодвигательного нерва (III пара), хорошо выраженное на 12-й неделе внутриутробного развития.
глазодвигательного нерва (III пара), хорошо выраженное на 12-й неделе внутриутробного развития.

 4.2. Ствол мозга
4.2. Ствол мозга


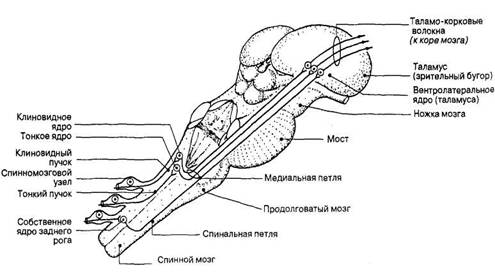
 срединной части покрышки правую и левую медиальные петли (lemniscus medialis). В продолговатом мозге к медиальной петле присоединяются волокна, восходящие из спинного мозга в составе спинальной петли, которая проводит импульсы, возникающие под влиянием тактильных, температурных и болевых воздействий. Таким образом, в составе медиальной петли проходят нервные волокна восходящего сенсорного пути, направляющиеся в промежуточный мозг (рис. 60).
срединной части покрышки правую и левую медиальные петли (lemniscus medialis). В продолговатом мозге к медиальной петле присоединяются волокна, восходящие из спинного мозга в составе спинальной петли, которая проводит импульсы, возникающие под влиянием тактильных, температурных и болевых воздействий. Таким образом, в составе медиальной петли проходят нервные волокна восходящего сенсорного пути, направляющиеся в промежуточный мозг (рис. 60).


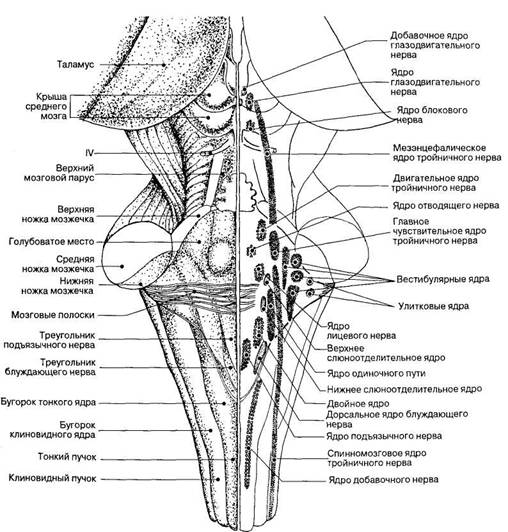
 В покрышке моста проходят волокна восходящих сенсорных путей (рис. 62). На уровне моста к медиальной петле присоединяются также волокна тройничной (тригеминальной) петли, образованной отростками вторых нейронов, лежащих в чувствительном ядре тройничного нерва. Эти волокна составляют восходящий сенсорный путь от головы (рис. 62). В покрышке моста проходят нервные волокна восходящего сенсорного пути, проводящего импульсы, возникающие вследствие проприоцептивных раздражений. Эти волокна образуют медиальную петлю. К медиальной петле присоединяются нервные волокна, идущие в составе спинальной петли. Нервные волокна спинальной петли проводят импульсы экстероцептивной чувствительности через мост к нервным центрам промежуточного мозга. Таким образом, нервные волокна, входящие в состав медиальной петли, спинальной петли и тригеминальной петли, которые объединяют под общим названием лемнисковые пути (lemniscus — петля), составляют важное звено сенсорных путей, проводящих сенсорную информацию в промежуточный мозг и далее в кору конечного мозга.
В покрышке моста проходят волокна восходящих сенсорных путей (рис. 62). На уровне моста к медиальной петле присоединяются также волокна тройничной (тригеминальной) петли, образованной отростками вторых нейронов, лежащих в чувствительном ядре тройничного нерва. Эти волокна составляют восходящий сенсорный путь от головы (рис. 62). В покрышке моста проходят нервные волокна восходящего сенсорного пути, проводящего импульсы, возникающие вследствие проприоцептивных раздражений. Эти волокна образуют медиальную петлю. К медиальной петле присоединяются нервные волокна, идущие в составе спинальной петли. Нервные волокна спинальной петли проводят импульсы экстероцептивной чувствительности через мост к нервным центрам промежуточного мозга. Таким образом, нервные волокна, входящие в состав медиальной петли, спинальной петли и тригеминальной петли, которые объединяют под общим названием лемнисковые пути (lemniscus — петля), составляют важное звено сенсорных путей, проводящих сенсорную информацию в промежуточный мозг и далее в кору конечного мозга.


 1 Здесь и далее употребляется латинизированное название покрышечно-спинномозгово-го пути (tractus tectospinalis), поскольку оно более точно отражает локализацию этого пути в стволе мозга.
1 Здесь и далее употребляется латинизированное название покрышечно-спинномозгово-го пути (tractus tectospinalis), поскольку оно более точно отражает локализацию этого пути в стволе мозга.

 ронах спинного мозга и симпатических преганглионарных нейронах его грудного отдела. Эти пути оказывают влияние на состояние спинномозговых мотонейронов, а также болевую чувствительность на уровне сегментарных связей.
ронах спинного мозга и симпатических преганглионарных нейронах его грудного отдела. Эти пути оказывают влияние на состояние спинномозговых мотонейронов, а также болевую чувствительность на уровне сегментарных связей.