|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Одновременно при применении Таи в гипоталамусеСтр 1 из 5Следующая ⇒
С. В книге представлены современные литературные данные, а также результаты собственных экспериментальных и клинических исследований по биохимии, фармакологии и медицинскому применению таурина (2-аминоэтансульфоновой кислоты) — продукта превращений серусодержащих аминокислот, относительно незаменимого нутриента и высокоактивного эндогенного регулятора метаболических процессов, эффективного средства целенаправленной коррекции и терапии широкого спектра патологических состояний, обладающее радиопротекторным, антиоксидантным, мембраностабилизирующим, нейроэффекторным, гепато- и кардиопротекторным действием на функциональные системы организма. Значительная часть представленных в книге результатов получена при выполнении заданий VI раздела Государственной научно-технической программы 43.01.р. "Создать новые эффективные лекарственные препараты на основе аминокислот и их производных" для их производства на Гродненском заводе медпрепаратов, в процессе реализации которых создан и зарегистрирован новый эффективный отечественный лекарственный препарат "таурин". Рассчитана на биохимиков, фармакологов, физиологов, медиков-практиков и экспериментаторов, а также студентов и аспирантов медико-биологического профиля. Табл. 35, Ил. 31, Библиогр.: 236 . Научный редактор академик И.Д.Волоховский Рецензенты: Чиркин А.А., д-р мед. наук, проф. Дубовик Б.В.., д-р мед. наук, проф. ОГЛАВЛЕНИЕ ВВЕДЕНИЕ 6 ГЛАВА 1. Основные физико - химические, токсикологические и фармакологические характеристики таурина 9 ГЛАВА 2. Биосинтез, транспорт и катаболизм таурина 17 ГЛАВА 3. Биологическая активность таурина 26 3.1. Мембраностабилизирующий и антиокидантный эффекты, 3.2. Влияние таурина на процессы формирования фонда свободных 3.2.1. Недостаточность таурина 75 3.2.1.1 Формирование фонда нейроактивных аминокислот и биогенных аминов в отделах головного мозга при недостаточности таурина 85 3.2.2. Избыточное поступление таурина 88 3.2.2.1 Однократное парентеральное введение 88 3.2.2.2 Длительное парентеральное введение 90 3.2.2.3 Длительное субконъюктивальное введение 97 3.3. Нейрэффекторное действие таурина и процессы формирования фонда 3.3.1 Парентеральное введение таурина 103 3.3.2. Субконъюнктивальное введение таурина 109 ГЛАВА 4. Клинические аспекты применения таурина 114 ГЛАВА 5. Механизмы регуляторных эффектов и стратегия использования L-аминокислот и их производных в качестве эффективных средств метаболической терапии и новых лекарственных препаратов 132 ЛИТЕРАТУРА 142 Принятые в тексте сокращения: ос-ААК (ос-ААА) а-аминоадипиновая кислота Р-Ала (р-А1а) р-аланин АЛТ (ALT) аланинаминотрансфераза (КФ 2.6.1.2)
а, -р-АМК (а, -р-АВА)а,-Р-аминомасляная кислота аминокислоты с разветвленной углеводородной цепью (валин, лейцин, изолейцин) аспартатаминотрансфераза (КФ 2.6.1.1) у-аминомасляная кислота З-метокси-4-оксифенилуксусная (гомованилиновая) кислота глутаматдегидрогеназа (КФ 1.4.1.2) глюкогенные аминокислоты 3,4-диоксифенилэтиламин (дофамин) 3,4-диоксифенилуксусная кислота заменимые аминокислоты изоцитратдегидрогеназа (КФ 1.1.1.42) кетогенные аминокислоты 2-оксоглутаровая кислота а-кетоглутаратдегидрогеназа(КФ.1.2.4.2.) малатдегидрогеназа (КФ 1.1.1.37) лактат (молочная кислота) мочевина З-метокси-4-оксифенилэтиленгликоль 3-метокситирамин незаменимые аминокислоты 5-оксииндолуксусная кислота 5-окситриптамин (серотонин) 5-окси-Ь-триптофан пируват (пировиноградная кислота) пируватдегидрогеназа (КФ.1.2.2.2.) серусодержащие аминокислоты L-триптофан центральная нервная система цистин цистатионин цитруллин фосфоэтаноламин этаноламин 1Ч-метил-р-(3,4-диоксифенил)-р-оксиэтиламин (адреналин) ароматические аминокислоты (фенилаланин, тирозин) восстановленный глутатион окисленный глутатион Р~(3,4-диоксифенил)-р-оксиэтиламин (норадреналин) аммиак S-аденозил-Ь-метионин SAH S-аденозил-Ь-гомоцистеин Таи (Таи) 2-аминоэтансульфоновая кислота (Таурин) т, с митохондриальная или цитоплазматическая форма ферментов ВВЕДЕНИЕ Таурин (Таи, 2-аминоэтансульфоновая кислота, H2N-CH2-CH2-SO3H) является конечным продуктом метаболизма серусодержащих аминокислот и их производных (глутатиона, метионина, цистеина, цистина) у млекопитающих. Кроме того, в организм Таи поступает с пищей животного происхождения и является для человека относительно незаменимым нутриентом [1]. Таи обладает радиопротекторным, антиоксидантным, мембраностабилизирующим действием, является тормозным нейромодулятором, проявляет гепато- и кардиопротекторные, антиаритмические и нормотензивные свойства [2-4]. Имеется достаточное число работ, демонстрирующих широкий спектр эффектов фармакологически активных доз этого соединения [5-9]. Однако до сих пор не исследованы регуляторные механизмы влияния Таи в концентрациях, сравнимых с эндогенными, на метаболические процессы, опосредующие реализацию его биологического действия, в частности, на процессы формирования фонда свободных аминокислот и родственных соединений, в том числе — биогенных аминов, на энергозависимые реакции промежуточного обмена, обмен нейромедиаторов. Невыясненными остаются также метаболические последствия недостаточности Таи в организме. Продемонстрировано, что эндогенная концентрация Таи является информативным показателем при патологических состояниях, сопровождающихся развитием метаболического дисбаланса (как клинических, так и воспроизведенных в эксперименте). Это позволяет предположить, что часть известных эффектов Таи опосредуется влиянием на процессы синтеза de novo и деградации свободных серусодержащих аминокислот, системы антиоксидантной защиты, функционирование транспортных систем, а также патогенетически обосновать рациональность его применения для коррекции метаболического дисбаланса [10]. Свойства этого соединения позволяют использовать лекарственные препараты на его основе как эффективные средства при поражениях печени [11-17], патологии сердечно-сосудистой системы [18-22], заболеваниях центральной нервной системы [23-25] катарактах и глаукоме [26], сахарном диабете [27], интоксикациях, а также наркоманиях и алкоголизме [28]. Кроме того, учитывая высокую степень нутриентной незаменимости этого соединения, оправдана необходимость введения Таи в состав продуктов питания, в первую очередь — детского [29-31]. Поэтому на сегодняшний день существует более чем достаточно предпосылок, обосновывающих актуальность фундаментальных исследований и прикладных разработок для дальнейших испытаний биологической активности и выяснению механизмов действия этого соединения, а также показаний и способов его применения в медицинской практике. Значимость таких исследований для народного хозяйства определяется их основной целью, заключающейся в определении научно обоснованных показаний и способов применения Таи для целенаправленной метаболической коррекции и терапии широкого спектра заболеваний и патологических состояний, включая прямо или косвенно связанные с воздействием неблагоприятных экологических факторов, в том числе ионизирующей радиации. Дальнейшее осмысление полученных результатов и развитие исследований в этом направлении требуют широкого обсуждения. В настоящей книге представлены современные литературные сведения, а также результаты собственных 15 летних экспериментальных и клинических исследований по биохимии, фармакологии и медицинскому применению Таи, ставшие основой для регистрации и внедрения в фармацевтическую промышленность нового эффективного отечественного лекарственного препарата гепато- и радиопротекторного действия "таблетки таурина", которому свойственны практически полное отсутствие побочных эффектов, возможность длительного приёма, усиления полезных эффектов других лекарств и препятствие проявлению их побочного действия, а также адаптогенные эффекты в отношении вредных факторов окружающей среды. Исследования в этой области выполнены в нами Институте биохимии НАН Беларуси в сотрудничестве с Институтом физико-органической химии НАНБ, а также с институтами МЗ РБ (Минский, Гродненский и Витебский мединституты) в рамках реализации заданий Государственной научно-технической программы "Лекарственные препараты". Поэтому изложенный материал представляет интерес не только для научных работников, но и практических врачей, специалистов в области технологии и производства лекарственных препаратов а также студентов и аспирантов медико-биологического профиля. ГЛАВА 1. ОСНОВНЫЕ ФИЗИКО - ХИМИЧЕСКИЕ, ТОКСИКОЛОГИЧЕСКИЕ И ФАРМАКОЛОГИЧЕСКИЕ ХАРАКТЕРИСТИКИ ТАУРИНА В данном разделе книги суммированы и анализируются литературные сведения, касающиеся основных токсикологических и фармакологических параметров высокое чищенной субстанции таурина (Таи), а также результаты собственных исследований, полученные в процессе изучения неспецифической фармакологической активности лекарственной формы "таблетки таурина" в рамках реализации заданий IV раздела Государственной научно-технической программы 43.01.р. "Создать новые эффективные лекарственные препараты на основе аминокислот и их производных" [32]. В настоящее время технология производства отечественного Таи освоена в Институте физико-органической химии НАН Беларуси, создана опытно-промышленная установка (содержание Таи в препарате не менее 99,0%), а промышленное производство нового лекарственного препарата "таблетки таурина" (зарегистрирован и разрешён к применению МЗ РБ решением Фармкомитета РБ от 25.06.97, N12-1-6/4271 в качестве гепато- и радиопротектора) на основе доступного сырья освоено на Гродненском (г. Скидель) заводе медицинских препаратов. В физическом отношении таурин (Таи, 2-аминоэтансульфоновая кислота, H2N — СН2 — СН2 — SO3H) представляет собой белый бесцветный кристаллический порошок с молекулярной массой 125,15 у.е. и температурой плавления 321-323°С (с разложением), без запаха, хорошо растворимый в воде (10,5г/100 мл при 25°С, прозрачный бесцветный раствор), 0,1М растворе HCL и практически нерастворимый в 95% этиловом спирте, эфире, ацетоне и хлороформе [1]. При исследовании токсикологических характеристик высокоочищенной субстанции Таи на мышах и крысах нами установлено, что вне зависимости от пола животных определение LD5o для неё затруднено. Так, при подкожном или внутрибрюшинном введении животным 8% раствора Таи LD50 для этого соединения составляет >4,0, а при пероральном введении 25% - 35% растворов Таи — от 6,50 до 7,23 г/кг. При этом назначаемый парентерально Таи вызывает минимальные изменения состояния и поведения животных, описываемые как снижение реакции на внешние раздражители, незначительно выраженные анорексию, гиподинамию и учащение дыхания. Поэтому в соответствии с классификацией СД.Заугольникова Таи относится к малотоксичным веществам [32]. Доказано также, что Таи не обладает способностью к кумуляции и его токсичность, оцениваемая по величине LD5o при многократном введении мышам, статистически значимо не отличается от таковой у животных, которым он назначался однократно, а расчётная величина индекса кумуляции (+2,25 — +3,6) свидетельствует о высокой степени обратимости токсического действия этого соединения [32]. При одно- или многократном закапывании в конъюнктивальный мешок 4% или 8% водного раствора Таи кроликам у них не отмечается блефароспазма, изменений в кровенаполнении сосудов роговицы, лакримации и повреждающего влияния на слизистую оболочку. Кроме того, в многочисленных клинических наблюдениях обосновано, что Таи не оказывает раздражающего действия на слизистую конъюнктивы человека и не изменяет величину зрачка, а в форме 4% раствора (лекарственная форма "Тауфон", Россия) Таи используется в виде глазных капель или для субконъюнктивальных инъекций в терапии офтальмологических заболеваний [1,26,32]. В экспериментах по исследованию общерезорбтивного действия субстанции Таи при длительном ежедневном эпикутанном назначении её 25% суспензии в дозе 1г/кг массы тела морских свинок показано, что это соединение обладает способностью всасываться неповрежденной кожей, поскольку при этом определяется значимое (в 1,5 раза) увеличение его концентрации в плазме крови экспериментальных животных. Тестируемые на этом фоне показатели раздражающего действия Таи на кожу, поведенческие реакции, объективный статус, массу тела, а также биохимические и морфологические параметры кожных покровов животных практически не отличались от таковых у интактных особей [32]. В испытаниях общей фармакологической активности Таи при его однократном назначении в дозах, равных 1/10 LD5o на кг массы тела животных и оценке через 30-60 минут показателей его нейротропного действия (таких, как спонтанной двигательной активности, ориентировочных реакций, эмоциональной реактивности, антиконфликтного эффекта, условных рефлексов, ректальной температуры, изменений координации движений, судорожной активности и развития максимального судорожного припадка) было продемонстрировано, что он подавляет у мышей спонтанную двигательную активность, оцениваемую по их горизонтальным перемещениям, угнетает ориентировочные реакции животных и оказывает гипотермическое действие, а при введении крысам проявляет антиконфликтные свойства и удлиняет время условного рефлекса активного избегания. После введения Таи в дозе 1/100 LD5o на кг массы животных описанные выше изменения не проявляются, а поведенческие реакции и ректальная температура у мышей и крыс не отличаются от исходного уровня [32]. В экспериментах по оценке эффектов Таи при его введении в дозе 1/10 LD5o на кг массы на эмоциональную реактивность мышей в соответствии с пяти-бальной шкалой Брэди и Наута показано, что на фоне стресс-провоцирующей стимуляции он индуцирует усиление эмоциональной реактивности животных, а при введении в дозе, равной 1/100 LD5o, не влияет на эти показатели. Одновременно, назначаемый в дозах 1/10 или 1/100 LD50 на кг массы Таи не изменяет координацию движений мышей, и их способность удерживаться на вращающемся стержне в течение контрольного времени. Кроме того, Таи при его предварительном, за 1 час до внутрижелудочного введения гексенала или хлоралгидрата, назначении мышам в дозах, равных 1/10 или 1/100 LD50, дозозависимо удлиняет продолжительность сна, усиливая снотворное действие указанных препаратов соответственно в 7 и 4 раза [32]. Показано также, что, вводимый в вышеуказанных дозах мышам, Таи подавляет судороги, вызываемые ареколином, не меняет интенсивность коразоловых судорог и не предупреждая на этом фоне гибели животных. Одновременно при введении в дозе 1/10 LD5o на кг массы, Таи снижает интенсивность судорог вызываемых никотином и предупреждает гибель животных, а при назначении в той же дозе за 1 час до введения тиосемикарбазида удлиняет латентный период индуцированных этим препаратом судорог и предупреждает гибель мышей, не препятствуя развитию тонико-экстензорной фазы судорожного припадка и гибели животных, развивающихся на фоне пропускания через корнеальные электроды переменного тока. Таким образом, Таи изменяет функциональные параметры центральной нервной системы, оказывая депримирующее действие и проявляя противосудорожные свойства [32]. В экспериментах по изучению действия Таи на показатели, характеризующие функциональное состояние сердечно-сосудистой системы, показано, что через короткие интервалы времени (1-60 минут) после его внутривенного введения в дозе 10-100 мг/кг крысам определяется дозозависимое повышение артериального давления главным образом за счёт увеличение ударного объёма. При этом на фоне компенсаторно возникающей брадикардии минутный объём и общее периферическое сопротивление практически не изменяются, а частота дыхания животных увеличивается на 40% [32]. При ежедневном внутрижелудочном введении Таи крысам с 1 по 6, с 6 по 16 и с 16 по 19 дни беременности крысам в дозах близкой к максимально переносимой (5,0г/кг), 1/10 LD50 для самок (0,72 г/кг) или дозе, близкой к расчётной для клинической апробации (1/100 LD5o - 72 мг/кг) доказано, что он не оказывает влияния на репродуктивную функцию и потомство животных, не обладает эмбриотропным (эмбриотоксическим, тератогенным и фетотоксическим) действием. Кроме того, продемонстрировано, что Таи назначаемый в вышеуказанных дозах, не влияет на плодовитость крыс, не изменяет количество жёлтых тел и мест имплантации, постимплантационную гибель плодов, индексы беременности и плодовитости самок, а при ежедневном энтеральном введении в дозе, равной 1/100 LD5o с 6 по 16 день беременности вызывает умеренное (5-7%), но достоверное увеличение средней массы плодов, плацент и плодно-плацентарного коэффициента, не оказывая влияния на количество родившихся живыми крысят и не изменяя показатели перинатальной смертности, физического развития потомства, а также скорости созревания сенсорно-двигательных рефлексов в период вскармливания новорожденных [32]. Одновременно у плодов не обнаруживается аномалий развития внутренних органов (сердца, лёгких, бронхов, пищевода, печени, кишечника, поджелудочной железы, мочеполовой системы), нижней челюсти, твердого неба, носовой полости, головного мозга, боковых, третьего и четвертого желудочков мозга и мозжечка), а назначение Таи в дозе 1/100 LD5o с 6 по 16 день беременности самок индуцирует увеличение массы тела крысят в течение их первой недели жизни [32]. Установлено также, что после сенсибилизации и последующей внутрикожной разрешающей инъекции 0,05 мл 0,01% раствора Таи у морских свинок и мышей регистрируются минимальные изменения внешних признаков поведения и практически не изменяется ректальная температура, что свидетельствуют об отсутствии у препарата анафи-лактогенной активности. Кроме того, на мышах-самках линии СВА показано, что Таи при его четырёхкратном подкожном введении через день в дозах 50 или 150 мг/кг после разрешающей инъекции не изменяет индекс воспалительной реакции и не влияния на реакцию гиперчувствительности замедленного типа [32]. В экспериментах по исследованию влияния Таи на гуморальный и клеточный иммунный ответ, основанного на определении титра агглютининов в сыворотке крови на 7,15 и 30 сутки после иммунизации мышей линии СВА эритроцитами барана, определении количества анти-телобразующих клеток на 5 сутки в селезенке, а также действии препарата на реакцию гиперчувствительности замедленного и немедленного типов, оцениваемом в опытах на морских свинках, сенсибилизированных подкожным введением 0,1мл лошадиной сыворотки, показано, что Таи практически не влияет на изученные показатели [32]. Хроническая токсичность высокоочищенной субстанции Таи оценивалась нами в экспериментах на крысах, кроликах и собаках при его ежедневном внутрижелудочном введении в форме взвеси в 2% крахмале в дозах 50мг/кг (близкая к расчетной суточной для клинического применения), 300 мг/кг (промежуточная) и 700 мг/кг (близкая к 1/10 LD5o) в течение 90 дней. Оказалось, что Таи, назначаемый в дозе 50 мг/кг не вызывает отрицательного действия на интегральные показатели объективного статуса и поведения животных (потребление пищи и воды, двигательную активность, реакции на внешние раздражители), а также на изменение массы тела. Одновременно показано, что у при назначении Таи в дозах 300 или 700мг/кг на фоне слабо выраженных гипотермии и брадикардии отмечается тенденция к учащению дыхания экспериментальных животных. Кроме того, Таи в указанных дозах практически не вызывает изменений в содержании эритроцитов и уровне гемоглобина, индуцирует незначительное уменьшение числа тромбоцитов в крови, а регистрируемый при этом на фоне повышения содержания лейкоцитов умеренный ретикулоцитоз свидетельствует об активной регенеративной способности костного мозга животных и отсутствии признаков угнетения гемопоэтической ткани. Общий анализ эффектов Таи на систему гемостаза позволяет сделать заключение о том, что он обладает антикоагулянтным действием. Об этом свидетельствует изменение параметров тромбоэластограммы (увеличение констант протромбиназы, тромбина, тотального свёртывания крови с одновременным уменьшением её максимальной амплитуды, константы эластичности сгустка, удлинения общего времени свёртывания крови), свидетельствующих об угнетении процессов образования тромбина и снижении концентрации фибриногена в крови. Указанные изменения обратимы, практически не зависят от дозы вводимого Таи, не носят прогрессирующего характера и не приводят к появлению кровоточивости [32]. Кроме того, хроническое назначение Таи в указанных дозах не изменяет диурез и концентрацию гиппуровой кислоты в моче, а, судя по показателям бромсульфалеинового теста, концентрациям мочевины и креатинина, не влияет на выделительную функцию печени и почек, обратимо увеличивая содержание общего белка, процентное содержание альбуминов и уменьшая относительный уровень глобулинов, особенно их OCi, OC2 и Р-фракций, снижая уровень общего билирубина и умеренно активируя активность процессов переаминирования в сыворотке крови. Кроме того, при хроническом назначении в дозе 300 мг/кг Таи оказывает обратимое липотропное действие, проявляющееся в снижении содержания общих липидов и их фракций в крови животных, а при его назначении в дозе, равной 700 мг/кг оказывает выраженный гиполипидемический эффект, оцениваемый по снижению содержания липидов, уменьшению концентраций общего холестерина, фосфолипидов, триглицеридов, (3-липопротеидов и неэстерифицированных жирных кислот. При длительном хроническом назначении Таи практически вне зависимости от назначаемой дозы и длительности введения и без выраженного влияния на активность лактатдегидрогеназы, а также уровни пирувата и лактата индуцирует умеренное обратимое снижение содержания глюкозы в сыворотке крови крыс, проявляя, таким образом, гипогликемические свойства [32]. Весовые коэффициенты головного мозга, лёгких, сердца и почек после хронического назначения Таи не изменяются, а для печени и селезенки крыс регистрируется их 15-20% увеличение. Морфологическое и гистологическое исследования органов и тканей крыс, получавших Таи в хронических экспериментах, не выявили деструктивных, дегенеративных и дистрофических изменений во внутренних органах [32]. Определение фармакокинетики вводимого внутривенно или орально в дозе 50 мг/кг меченного 14С-Таи мышам с определением методом нелинейной регрессии параметров двухкамерной модели показало, что динамика средних концентраций препарата в крови (рис.1) описывается приводимым ниже биэкспоненциальным уравнением:
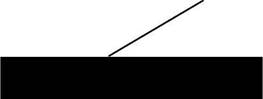
Ct=64,lez'36t+10,9e -0,112t (1) Из результатов, представленных на рисунке 1 и в таблице 1, видно, что период полуэлиминации Таи составляет 6,16 час, он быстро распределяется по органам и тканям со средним периодом полураспределения 0.29 час. При этом константа равновесного распределения составляет 3,3, а общий кинетический объем распределения в 4,3 раза больше объема в центральной камере (Vi), что подтверждает интенсивный захват Таи органам и тканями из крови [33] \ £ D Рис.1. Динамика средних концентраций С-Tau, вводимого внутривенно или орально (50 мг/кг) у мышей Фармакокинетические параметры таурина Таблица 1
Показатель
Период полуэлиминации (tm), ч Комплексный параметр для (3-фазы, ч"1 Период полураспределения (ti/2, а), ч Константа скорости переноса из цент-ральной камеры в периферическую 1.44 Константа скорости элиминации (Кэл), ч -1 0.61 Общий клиренс (Clt), л/ч 0.014 Объем распределения в центр, камере (Vi), л 0.023 Стационарный объем распределения (Vss), л 0.0995 Площадь под кривой (АИС) при введении в вену (АИСуу) и в желудок 123.8 (АИСр/„), мкМ/г 89.9 Абсолютная биодоступность, % 72.6 Константа скорости всасывания (Koi), ч"1 19.8 Период полувсасывания, ч 0.035
П — Ч1 К -I77t , г г -0,09t r>j i -19,8t /4,4 Ct=51,oe + 5,5 e — 57,1 e (2) Из полученных результатов следует, что фармакокинетические параметры 14С-Таи, определённые для орального способа введения, во многом схожи с таковыми при внутривенном способе его назначения. При этом константа скорости всасывания Таи из желудочно-кишечного тракта очень высока (K0i=19,8 ч'1), а время достижения максимума концентрации соединения в крови (tmax) составляет 0,035 часа. Степень биодоступности Таи при его пероральном введении, рассчитанная по величинам площадей под кривыми концентрации интегрированием уравнения 2 при равновесии доз составляет 72,6%, т.е. в системный кровоток всасывается около 2/3 Таи. При этом наибольшие количества меченого Таи уже через 10 минут регистрируются в печени и почках с последующим экспоненциальным снижением его концентрации в этих органах, сходным с таковым для крови. Кроме того, оказалось, что в достаточно больших количествах 14С-Таи определяется в сердечной мышце (пик концентрации на 45 мин), гораздо ниже его содержание в головном мозгу, что предполагает относительно плохое проникновение соединения через гематоэнцефалический барьер [32]. Полученные нами данные по экскреции радиоактивного Таи с мо-чой после его внутривенного, подкожного или внутрижелудочного введения свидетельствуют о том, что он быстро экскретируется. Так, уже через 3 часа после внутривенного введения в моче определяется 68,8%, а при подкожном или пероральном — соответственно 63,3 и 54,4% метки. Одновременно при внутривенном введении 14С-Таи через 24 часа с калом выделяется 6,6%, а при назначении через рот - 4,5% метки [32]. Расчёт экскреции 14С-Таи в течение 12 часов при различных способах его введения и определение его биодоступности у крыс и мышей показали, что при назначении через рот последняя составляет в среднем 74,2%, а при подкожном введении - 94,5%. Таким образом, литературные данные [1] и результаты проведенных нами токсикологическихи фармакологических исследований неспецифической активности высокоочищенной субстанции Таи доказывают, что он мало токсичен для лабораторных животных, не куму-лирует в организме, не обладает раздражающим действием на кожу и слизистые, не влияет на репродуктивную функцию и потомство крыс, не проявляет аллергизирующих свойств и не оказывает отрицательного влияния на иммунитет, а при хроническом его назначении возникающие изменения морфологических, физиологических и биохимических показателей не носят повреждающего характера, обратимы и возвращаются к исходному уровню после отмены препарата в течение 8-10 суток. Так, максимальная доза Таи, испытанная в клинике и не вызвавшая токсических проявлений — 15 г в день [1,23,24]. ГЛАВА 2. БИОСИНТЕЗ, ТРАНСПОРТ И КАТАБОЛИЗМ ТАУРИНА В животных клетках, по сравнению с растительными, Таи находят в значительных количествах. Он не инкорпорируется в белки и его концентрация во многих тканях достаточно велика: наибольшее содержание его характерно для тканей сердца (10-40 цмоль/г), головного мозга (2-6 цмоль/г), скелетных мышц (2-15 ц,моль/г), где им в зависимости от вида млекопитающих представлено от 30 до 50% всего аминокислотного фонда. Так, например, у крыс Таи составляет 0,15% от общей массы тела, что соответствует его концентрации 1200 р.моль/кг массы тела животных, а 75% всего пула Таи сконцентрировано в мышцах и сердце [34]. В головном мозге Таи — одна из пяти количественно преобладающих аминокислот (наряду с глутаматом, глутамином, ГАМК, глицином), где его концентрация видоспецифична, а распределение относительно монотонно. Большая часть его при этом сосредоточена в глие. Уровень Таи в синаптических пузырьках составляет 1% от общего содержания в головном мозге и, возможно, представляет собой нейротрансмиттерный пул [5]. Однако, пока нет данных о наличии Таи в синапсах. Кроме того, для мозга характерно эмбриональное накопление и постепенное снижение его концентрации вдвое в постнатальном периоде, хотя общий пул соединения с увеличением массы органа растет. На этом основании Таи называют "фактором роста мозга" [25, 35]. У эмбрионов и взрослых особей уровни Таи и глутамата, особенно в нервных окончаниях, отрицательно коррелируют, и снижение концентрации первого сопровождается увеличением второго [36]. Содержание крыс в течение трёх месяцев на диете с 3Н-Таи и 358-метионином показало, что 58% Таи поступает с пищей, 29% синтезируется de novo, a 13% представлено остаточным пулом тканей. При исключении Таи из диеты синтез его возрастает до 54%, а остаточный пул — до 46%. Адаптивные механизмы поддержания уровня Таи в тканях, таким образом, реализуются за счёт не только активации его синтеза, но и реабсорбции в почках [37,38]. Процесс биосинтеза Таи является одним из важнейших в катаболизме серусодержащих аминокислот у большинства видов млекопитающих. Скорость синтеза варьирует между видами: она относительно высока у крыс по отношению к человеку, тогда как кошки полностью лишены способности синтезировать Таи [4]. У крыс для его синтеза используется 25% серусодержащих аминокислот, а из двух основных путей распада цистеина в печени (до пирувата или Таи) "тауриновый" составляет 70%. У людей Таи занимает значительно меньшее место в катаболизме серусодержащих соединений. Так, отношение экскретируемых сульфатов к Таи у человека составляет 45, а у крыс и мышей — 3 [34, 39]. В организме млекопитающих существует быстро обмениваемый (с периодом полураспада 2-4 дня) и медленно обмениваемый (с периодом полураспада 20-30 дней) пулы свободного Таи [40]. Быстро обмениваемый пул формируется за счёт пищевого Таи и его синтеза de novo. Небольшая часть этого пула пополняет медленно обмениваемый, используется для конъюгации с желчными кислотами, экскретируется с мочой и калом в виде конечных продуктов распада, образующихся под действием микрофлоры кишечника. Желчный Таи реабсорбируется в кишечнике и почках [3]. По скорости оборота Таи ткани млекопитающих подразделяются на 3 группы: первая — с высокой скоростью оборота (печень, почки, поджелудочная железа, надпочечники), вторая — с умеренной скоростью (легкие, селезенка, кишечник) и третья — с малой скоростью (сердце, мышцы, мозг). Уровень удельной радиоактивности введенного внутрибрюшинно 35S-Tau достигает максимума в первой группе тканей через 1 сутки после инъекции, во второй — на 2-3 сутки, а в третьей — на 4-7 сутки [40]. Период полураспада Таи у мышей, определенный по включению 358-метионина, равняется 18,6 дня. У крыс он составляет 11,4 дня и при содержании их на бесТауриновой диете увеличивается до 15 дней [39]. Основным предшественником Таи является поступающий с пищей или формирующийся из метионина цистеин (рис. 2). Печень крысы за сутки окисляет около 400 цмоль цистеина до Таи. Экзогенный цистин, восстанавливаясь перед его проникновением в клетку глутатионредуктазной системой до цистеина, также вносит важный вклад в пул предшественников биосинтеза Таи. У крыс за сутки на 100 г массы тела синтезируется до 35 цмоль Таи, т.е. для этого используется 15-30% серы из метионина и цистеина (рис.2). При интрагастральной нагрузке цистеином из него образуется гораздо меньше (45%) Таи, чем при интраперитонеальной (83%). Это объясняется способностью клеток кишечника метаболизировать цистеин до пирувата через активацию процессов трансаминирования [37,41]. Хотя использование цистеина для образования Таи весьма значительно, обсуждение его роли в поддержании "тауринового" гомеостаза требует учёта процессов трансаминирования, активно протекающих в печени, сердце и почках. Так, с учетом вклада трансаминирования промежуточных продуктов на этапах синтеза Таи из экзогенного цистеина in vivo соотношение путей утилизации последнего до пирувата или Таи в печени становится равным. Цистеин может активно трансаминироваться с 2-оксоглутаратом или пируватом с образованием меркаптопирувата, глутаминовой кислоты и аланина (рис.2) [34,39]. Из трёх ферментов катаболизма цистеина (цистеин -2-оксоглутаратаминотрансфераза, цистеин -пируватаминотрансфераза и 3-меркаптопируватсульфотрансфераза) первые два наиболее активны в сердце, а последний — в печени, почках и крови. Трансаминирование цистеина происходит преимущественно в митохондриях, а десульфурирование равномерно распределено между последними и цитоплазмой [42-45]. Процесс переаминирования цистеина стимулируется при добавлении в реакционную среду пирувата и ос-кетоглутарата [46, 47]. Первый этап окисления цистеина до Таи осуществляется под действием цистеин диоксигеназы (КФ 1.13.11.20) [47-50]. Км для цистеина в этой реакции (печень крыс) значительно меньше, чем в цистеин-трансаминазной (КФ 2.6.1.3) реакции. Поэтому трансаминазный путь деградации цистеина становится особенно значимым при увеличении концентрации субстрата [119,143]. Фермент очень активен в печени, мозге, сердце, селезенке и локализован преимущественно в цитоплазме [5,39,51]. Как и для целого ряда ферментов метаболизма серусодержащих аминокислот, для него характерна постнатальная активация. Полупериод его деградации в печени составляет 3,7 ч и, в отличие от других ферментов деградации цистеина, не реагирующих на избыток субстрата, увеличивается почти в два раза при парентеральной нагрузке цистеином [49, 52]. Цистеин-оксигеназа in vivo в значительной мере может активироваться гидрокортизоном и NAD+, a in vitro ингибируется кислородом, гомоцистеином и Са2+ [53]. Субстратная индукция фермента из печени крыс, осуществляемая за счет его синтеза de novo, аддитивно усиливается NAD+ или гидрокортизоном [54]. Образующийся в процессе окисления цистеина важнейший предшественник Таи цистеинсульфинат под действием митохондриальной дегидрогеназы печени в незначительной степени превращается в цистеат [55]. В сердце, почках и печени оба продукта могут переаминироваться с пируватом и 2-оксоглутаратом [46]. У крыс 20% образующегося в печени цистеинсульфината трансаминируется до Р-сульфинилпирувата, распадающегося до пирувата и сульфита спонтанно или под действием микрофлоры кишечника [56, 57]. Избыток цистеина в диете не влияет на активность переаминирования цистеинсульфината с 2-оксоглутаратом [37, 58]. Для мозга крыс показано, что цистеинсульфинаттрансаминаза и аспартатаминотрансфераза являются изоферментами с молекулярными массами 84 и 86 КД соответственно и состоят из двух субъединиц. В митохондриях выше активность первого изоэнзима, в цитозоле — второго. Км для 2-оксоглутарата у этих ферментов составляет соответственно 3-Ю"4 М и 1,5-10"4 М. Одновременно, концентрации аспартата и цистеинсульфината в организме гораздо ниже, чем 2-оксоглутарата [37]. В мозге активность его трансаминирования на два порядка выше, чем декарбоксилирования [5]. Приведенные данные подтверждают важность процессов трансаминирования в метаболизме серусодержащих аминокислот и их прямую причастность к образованию Таи. Цистеинсульфинатдекарбоксилаза (КФ 4.1.1.29) — пиридоксальзависимый цитозольный фермент, субстратами для которого могут служить как цистеинсульфинат, так и цистеат. In vitro он реактивируется при внесении кофактора (пиридоксаль-5-фосфат-эффект), a in vivo очень чувствителен к недостатку пиридоксина: В6-дефицитная диета вызывает снижение активности фермента уже через несколько дней. В печени крыс 80% цистеинсульфината декарбоксилируется до гипотаурина (2-аминоэтансульфинат) [34, 56, 59]. Декарбоксилазная активность по отношению к цистеинсульфинату в 6-8 раз выше, чем к цистеату, что показано на примере печени крыс, мышей и собак [39]. Км для обоих субстратов равняется 1,4-10'4М и 4,0-10' 4 М соответственно. Активность декарбоксилазы в печени с цистеинсульфинатом достигает 45, а с цистеатом — 5-10 цмоль/г-ч. Активность фермента видоспецифична и слабо коррелирует с концентрацией Таи и его обменом в тканях крыс. В печени человека фермент в 100 раз менее активен, чем у крыс [34]. В отличие от цистеин-оксидазы, которая активируется в печени крыс избытком экзогенного метионина и цистеина, цистеинсульфинатдекарбоксилаза наполовину ингибируется в этих условиях. У самцов активность этого фермента выше, чем у самок, а инъекции эстрадиола вызывают резкое его ингибирование.
aeiidaddei aaaeadiaaiaga dendaei aaaeadiaai aga \ dendaei пдёйдёшд ааёадаШпёёада Рис.2 Схема биосинтеза таурина (по A.Timbrell et.al [41]). Обозначения: ПАЛФ — пиридоксаль-5-фосфат, ФАФС — фосфоаденозилфосфосульфат, SAH — S-аденозилгомоцистеин, SAM — S-аденозилметионин. Введение кортизона, наоборот, активирует фермент в печени. Наиболее активен фермент в печени, мозге, почках, а наименее — в сердце. В результате более низкой активности по сравнению с цистеин-оксидазой, декарбоксилаза может лимитировать синтез Таи в мозге [37]. В отличие от печеночной, цистеинсульфинатдекарбоксилаза в мозге менее вариабельна по активности у различных видов млекопитающих, а по своей локализации в клетке и свойствам схожа с глутаматдекарбоксилазой [35]. Глутамат in vitro ингибирует образование Таи, а высокие концентрации Таи в коре головного мозга всегда ассоциированы с низким уровнем глутамата. Глутаматдекарбоксилаза из мозга быка катализирует образование из цистеата или цистеинсульфината Таи или гипотаурина [5, 25]. Фермент, ответственный за превращение в печени гипотаурина в Таи, не выделен, что, вероятно, говорит о существовании в печени механизмов быстрой утилизации промежуточного продукта (гипотаурин определяется только в регенерирующей печени, видимо, из-за превалирования процесса окисления цистеинсульфината над декарбоксилированием). Известно лишь, что это процесс энзиматический, зависит от никотинамидных коферментов, in vivo активно протекает в печени, мозге и почках, a in vitro практически отсутствует. В печени крыс обнаружена гипотауринаминотрансфераза, имеющая Км по отношению к гипотаурину 8-10"3М, а к пирувату — 1-10"3М [38, 60]. Активность этого фермента в регенерирующей после частичной гепатэктомии печени крыс не меняется [36,39]. Для сердечной мышцы свойственна очень низкая цистеинсульфинат- и цистеатдекарбоксилазная активность. В опытах in vivo показано, что в миокарде всего 4-5% цистеина, добавленного в среду инкубации, превращается в Таи. Наличие наряду с этим высоких концентраций Таи в сердце дало повод для поиска альтернативных источников его образования. Им в первую очередь стал впоследствии достаточно убедительно обоснованный цистеаминовый путь [39]. Гипотаурин под действием высокоспецифичной оксидазы может окислятся (90%) до Таи или переаминироваться (10%) до сульфинацетальдегида [38]. Более сложным является вопрос о синтезе самого цистеамина. Фармакологические аспекты действия последнего (радиопротекторное, антиоксидантное) изучены более детально, чем пути превращений. Показано, что он является естественным продуктом обмена веществ у млекопитающих: может образовываться из меченых предшественников (метионина и цистеина) или в результате гидролиза ацетил-КоА и экскретируется с мочой. Специфичной для цистеина декарбоксилазы не найдено, а в сердце крыс для образования цистеамина расходуется около 4% ацетил-КоА [61]. Гипотетически предполагают даже, что цистеамин может образовываться при транссульфулировании метионина до цистеина [62]. Подобно другим аминотиолам он обладает способностью образовывать дисульфиды с SH-группами белков, и большая часть его находится в связанном состоянии. Поэтому истинная концентрация цистеамина в тканях, вероятно, в 30-40 раз превосходит определяемую (1 и 4-10 нмоль/г для печени и сердца соответственно) [63]. Одним из наиболее реальных цистеаминсодержащих продуктов является пантотеин, производное которого — 4-фосфопантотеин — субстрат двух метаболических (до дефосфо-КоА или пантотената) потоков, регулируемых отдельно через фосфатазную или пантотеиназную реакции. Его деградация — своеобразный челнок для активации декарбоксилирования цистеина. Гидролиз с образованием цистеамина в организме животных — единственная специфическая реакция деградации Ко А и производных пантотената [64]. Определенный вклад в тауриновый пул сердца, кроме того, вносит сериндегидратаза (КФ 4.2.1.13) с последующим сульфурированием образующегося из серина а-аминоакрилата фосфоаденозин-фосфосульфаттрансферазой (КФ 2.3.2.-.) до цистеата [34]. Количественно он не охарактеризован и является своеобразным шунтом транссульфурирования: сера в данном случае не переносится, как обычно, с цистеина или метионина на углеродный скелет серина, а активируется сульфурилазой до аденозин-фосфосульфата, который в киназной реакции фосфорилируется до фосфоаденозинфосфосульфата. Этот источник синтеза Таи доказан не только для сердца, но и для печени млекопитающих, где его значимость возрастает в случаях ограничения возможности синтеза Таи по другим путям [38]. Проблематичным пока остается существование наработки Таи in vivo по пути цистин — цистиндисульфоксид — цистеамин-дисульфоксид — гипоТаурин — Таи. Возможность такой последовательности превращений доказывается формированием цистеаминдисульфоксида in vitro и экскреции его, наряду с гипотаурином, с мочой при скармливании крысам цистеина. При даче цистиндисульфоксида с пищей собакам он определяется в печени, а экскреция Таирохолатов с калом при этом повышается [34,39]. Менее существенным источником эндогенного Таи у млекопитающих можно считать синтез его микрофлорой кишечника из сульфита или сульфата [34]. Na+-3aBHCHMbie транспортные системы Р-аминокислот, участвующие в транспорте Таи и р-Ala, выделены из мембран ретины, почек, саркоплазматических, синаптосомальных и других плазматических мембран [65]. Км для транспорта Таи из плазмы в эритроциты составляет 1-Ю'4 М, процесс поступления его в клетку активируется ионами Na+ и подавляется в анаэробных условиях [5, 66]. В эпителии кишечника крыс выделена №+-зависимая транспортная система для Таи с Км=2,5-10'4 М; транспорт в клетки осуществляется против градиента концентрации за счет трансмембранного градиента Na+, СГ и мембранного потенциала [65]. В клетках гепатоцитов показано наличие раздельного механизма для захвата и выхода Таи, причем захват, осуществляемый двумя насыщаемыми транспортными системами с Км= 8,9-10"2 и 4,47-10"3М соответственно, ингибируется другими Р -аминокислотами, а выход значительно медленнее и не зависит от концентрации Na+ [67]. Введение 3% раствора Р-аланина — структурного аналога Таи — вызывает снижение транспорта последнего в различные ткани (особенно печень) у мышей и увеличение его экскреции с мочой [68]. Гипотаурин конкурентно ингибирует транспорт Таи в клетку, а цистеат не влияет на этот процесс [69]. При высоком содержании Таи в пище скорость его транспорта в изолированных сегментах почек снижается, а адаптация к высокой концентрации Таи в пищевых продуктах проявляется у крыс уже через три дня после начала измененного питания и заканчивается на 7-10-й день. Отношение концентрации Таи в полости почечных канальцев к его содержанию во внеклеточном пространстве равно 25 и подтверждает наличие активного переноса Таи в клетку [70]. Концентрационный градиент для Таи по отношению к миокардиоцитам равен 400:1. Насыщаемость, специфичность и ингибирование структурными аналогами доказывает присутствие в сердце активных транспортных систем для Таи [39]. На изолированном сердце показано, что перенос Таи в клетки миокарда осуществляется с образованием промежуточного комплекса с переносчиком, удаление же К+ или Na+ из среды инкубации на порядок снижает поглощение его миокардом [71]. Содержание Таи в диете не влияет на его концентрацию в плазме крови, что в некоторой степени говорит о высокой приспособляемости его транспортных систем [3, 5, 40]. Значительная часть экзогенного Таи экскретируется с мочой: получая перорально Таи, интактные крысы экскретируют около 30% поступившего с пищей соединения, а в суточной моче хорьков, которым парентерально инъецировали 14С-Таи, содержится 26% от введенной дозы. Снижение концентрации Таи в тканях сопровождается усилением его реабсорбции в почках и уменьшением экскреции с мочой [38, 72]. Экскреция Таи с мочой видоспецифична, повышается при несбалансированном белковом питании, введении цистеина, кортикостероидов, на фоне облучения, алкогольной интоксикации, мышечной дистрофии, при гепатитах, нефритах [34]. Процент тауро-конъюгатов с холевыми кислотами от общей суммы тауро- и гликохолатов у млекопитающих видоспецифичен — от 5% у людей и обезьян до 80% у крыс [1]. После поступления желчи в кишечник и последующего его гидролиза Таи разлагается кишечной микрофлорой до сульфата. Это важный путь его элиминации из организма [3]. Превращение Таи до карбомоил-таурина, тауро- и фосфотауроциамина, определяемых в тканевых экстрактах и моче млекопитающих незначительно. Утилизация его до изотионата под действием тауриндегидрогеназы через промежуточную стадию образования в сульфацетальдегиддегидрогеназной реакции сульфацетальдегида осуществляется, как показано на крысах-гнотобионтах, лишь под действием кишечной микрофлоры [73]. ГЛАВА 3. БИОЛОГИЧЕСКАЯ АКТИВНОСТЬ ТАУРИНА В этом разделе суммированы и анализируютя современные литературные сведения и результаты собственных исследований многочисленных биологических эффектов и механизмов реализации метаболической активности Таи (рис.3) 3.1. Мембраностабилизирующий и антиокидантный эффекты, антиатерогенное, кардио-, радио- и гепатопротекторное действие В настоящее время доказано, что отдельные эффекты Таи связаны с его участием в окислительно-восстановительных реакциях. Так, например, продемонстрировано, что в культуре клеток (нейтрофилы и эозинофилы) он хлорируется, нейтрализуя ПОС1, продуцируемую миелопероксидазой, формируя стабильные хлорамины и тем самым предотвращая клетки от аутолиза. Кроме того, показано, что благодаря способности реагировать с гипохлоратом (прооксидантом) и удалять его с образованием относительно стабильного таурохлорамина, Таи ослабляет повреждения ДНК, вызванные ароматическими аминосоединениями in vitro. Высокий уровень Таи и гипотаурина обнаружен также в половых железах, где предполагается, что последний, окисляясь до Таи, действует как антиоксид ант [9, 74].
I
ДТаё TT^a Iueou Пабаоа Пса пао^аоёа аёаса
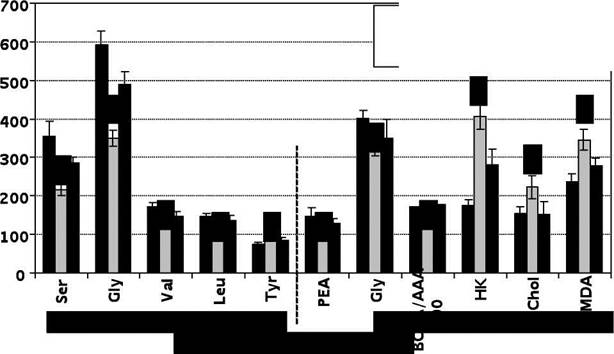
Рис.3 Основные функции и содержание таурина в организме млекопитающих Показано также, что in vitro Tau препятствует повреждению клеточных мембран различных тканей и клеток (скелетных мышц, миокарда, эритроцитов) внешними агентами, защищает сетчатку глаза и сперматозоиды от действия окислителей, гепатоциты — от токсического влияния CCLt, благодаря гипероксигенации подавляет перекисное окисление липидов, стабилизирует мембранную проницаемость и транспорт ионов [2, 8, 75-77]. Таи способен также предотвращать изменения внутриклеточого объёма, вызванные изменением осмолярности плазмы. Это можно связать со его способностью модулировать уровень Са2+ посредством понижения выхода последнего из митохондрий. Предполагается также, что Таи таким же путём, как с фосфатидилэтаноламином, может взаимодействовать с некоторыми мембранными белками, например, с фосфолипид-метилтрансферазой, нарушая структуру белков мембраны и её липидной подложки. Это, в свою очередь, может приводить к изменению функций мембраны. Следовательно, этот тип изменений в функции клеточной мембраны, или другие, например, мембранного потенциала, может привести к изменению внутриклеточной концентрации ионов (Са2+ и др.), а способность модулировать их клеточную концентрацию и, тем самым, регулировать объём клетки, может лежать в основе протекторных свойств Таи [41]. С мембраностабилизирующей функцией Таи связывают и его нейроэффекторное действие, которое подробно обсуждается нами ниже (гл.3.3), предположительно связано с модуляцией ионных потоков, изменением в мозге активности пируватдегидрогеназы, концентрации глутамата, а также подавлением высвобождения возбуждающих нейротрансмиттеров [6,39, 78, 79]. Антиоксидантные и мембраностабилизирующие свойства Таи, предполагающие его антиатерогенное действие, проявляются в меньшей степени, чем у типичных представителей этой группы соединений, однако следует учесть, что Таи содержится в организме в относительно высоких концентрациях и может назначаться в больших дозах [1]. Известно также, что Таи, наряду с глицином, участвует в образовании в печени первичных желчных кислот: глико- и таурохолатов. Синтез последних филогенетически сформировался раньше, чем гликохолатов [34]. Их присутствие способствует абсорбции липидов, липолизу, всасыванию жирных кислот в кишечнике. С другой стороны, конъюгация Таи с желчными кислотами влияет на элиминацию холестерина из организма и тем самым контролирует холестерогенез [80, 81]. Показано, что назначение Таи крысам, кошкам, индюшатам и цыплятам (1% раствор с пищей или с питьевой водой) активирует процессы конъюгации желчных кислот и увеличивает секрецию желчи [83], а при содержании крыс на высокожировой диете с добавкой Таи последний подавляет подъем холестерина в печени, ингибируя его кишечную абсорбцию [83, 84]. Кроме того, введённый в дозах 250 и 500 мг/кг массы тела Таи активирует транспорт холестерина из крови и его метаболизм до желчных кислот, а его конъюгация с холатами усиливает элиминацию холестерина и желчных кислот из организма с фекалиями, активирует транспорт холестерина из крови и его метаболизм до желчных кислот [85]. Особенно выражено гипохолестеринемическое действие Таи на фоне потребления высокожировой диеты с избытком холестерина [86,87]. Подобные результаты получены в опытах на морских свинках и крысах с экспериментальным холестазом [88]. Экскреция Таи происходит в основном с желчью. Он конъюгирует как с желчными кислотами, так и с ксенобиотиками (чаще — с органическими кислотами) [85]. При даче в течение 2-5 дней 0,5% раствора Таи морским свинкам увеличивается секреция желчи и замена в конъюгатах с желчными кислотами глицина на Таи, уменьшается индуцированный монооксижелчными кислотами холестаз [34]. Добавление 300-500мг Таи в диету снижает концентрацию желчных кислот и холестерина в желчи обезьян и усиливает синтез таурохолатов у поросят [7, 89]. Возможно, что высокий уровень таурохолатов у некоторых видов млекопитающих (например крыс) затрудняет моделирование экспериментального атеросклероза, т.к. скорость обмена желчных кислот увеличивается благодаря образованию холилтаурина. Через образование тауро-конъюгатов с желчными кислотами осуществляется взаимосвязь стероидогенеза и метаболизма серусодержащих аминокислот [90-92]. Антиатерогенные свойства Таи могут быть также обусловлены его регуляторным действием на ключевые реакции промежуточного обмена серусодержащих аминокислот и в целом процессы формирования внутриклеточного фонда свободных аминокислот и их дериватов — биогенных аминов [5,15, 23, 24, 79, 93-95]. Кроме того, как показано в ряде исследований, высокие концентрации Таи в тромбоцитах определяют участие этого соединения в процессах агрегации, тромбирования артерий и регуляции сосудистого тонуса [5, 96]. Антиатерогенное действие Таи подтверждено нами в модельной ситуации экспериментального атеросклероза у крыс-самцов линии Wistar CRL:(WI) WU BR, содержавшихся в течение 45 суток на атерогенной диете. Адекватность экспериментальной модели подтверждена нами патоморфологической характеристикой специфических изменений эндотелия сосудов [96]. Кроме того, судя по значительному повышению уровней холестерина в печени и плазме крови опытных животных, а также концентрации малонового диальдегида, активности метаболических реакций (гексокиназа, АсТ, АлТ) и содержанию субстратов (лактата и пирувата) в печени, нами in vivo смоделирована адекватная атеросклерозу метаболическая ситуация [97,98]. В описанных условиях дополнительное парентеральное введение Таи содержавшимся на атерогенной диете крысам индуцировало патоморфологическую и биохимическую регрессию присущих атеросклерозу признаков: содержание холестерина в печени и плазме крови и малонового диальдегида в печени снизилось до контрольных показателей у интактных животных, практически не отличалась от нормальных значений и активность гексокиназы, повышенная более чем в 2 раза в печени животных опытной группы (рис.4), что подтверждает наличие у Таи антиатерогенных свойств.
Рис. 4 Концентрации свободных аминокислот и их производных в плазме крови (мкМ), печени (мкмоль/г хЮО), активность гексокиназы (нмоль/г/мин хЮО), концентрации холестерина (мг% хЮО) и малонового диальдегида (нмоль/г хЮО) в печени крыс на фоне экспериментального атеросклероза и введения таурина, 650 мг/кг Это утверждение согласуется и с полученными нами результатами по исследованию пула свободных аминокислот в плазме крови и печени находившихся на атерогенной диете крыс, потребление которой вызвало снижение содержания АРУЦ и повышение концентрации тирозина в плазме крови, что указывает на снижение антитоксического индекса печени (так называемый индекс Фишера) [99]. При этом отмечался также явный дефицит гликогенных (глутамата, серина, глицина) аминокислот (рис.4). После введения Таи концентрации серина, глицина, валина, лейцина и тирозина в плазме крови возвращались к контрольным значениям (рис.4), что указывает на его способность нормализовать величину индекса Фишера, а также восстанавливать при атеросклерозе индуцированные торможениим ключевых реакций глюконеогенеза изменения в фонде гликогенных аминокислот [100]. Кроме того, в печени животных на фоне экспериментального атеросклероза повышается содержание глутамата, аланина и снижаются уровни глицина, этаноламина, фосфоэтаноламина и аммиака (рис.4). Само по себе снижение содержания этаноламина может свидетельствовать либо о нарушении деградации его предшественника серина, либо о снижении антиоксидантной защиты в печени [101,102]. С учётом однонаправленености сдвигов в содержании этаноламина и АРУЦ при неизменных концентрациях ароматических аминокислот последнее предположение является наиболее вероятным, а повышение, в свою очередь, концентраций глутамата и аланина в печени крыс, находящихся на атерогенной диете, может свидетельствовать об ингибировании глюконеогенеза [103]. Введение Таи на фоне экспериментального атеросклеорза приводило к повышению этого соединения в печени, с одновременным увеличением в ней концентраций лейцина, изолейцина и цистина и нормализацией содержания фосфоэтаноламина (рис.4). Таким образом, при эксеприментальном атеросклерозе Таи обладет способностью уменьшать проявления метаболического дисбаланса в плазме крови и печени, включая нормализацию фонда свободных аминокислот и их дериватов (увеличение уровня фосфоэтаноламина, соотношения АРУЦ и ароматических аминокислот). Исследование закономерностей формирования фонда нейроактивных соединений в отделах головного мозга показало, что введение Таи на фоне атеросклероза вызывало повышение содержания ГАМК в гипоталамусе при неизменном относительно опытной группы атеросклероза содержании "возбуждающих" медиаторов (Asp и Glu), которые были достоверно повышены в среднем мозге животных (рис.5). Последнее может свидетельствовать о нормализующем действии Таи на соотношение уровней "тормозных" и "возбуждающих" медиаторов в головном мозге при атеросклерозе, одним из звеньев патогенеза которого считают нарушения функционалой активности ГАМК-ергической системы [104].
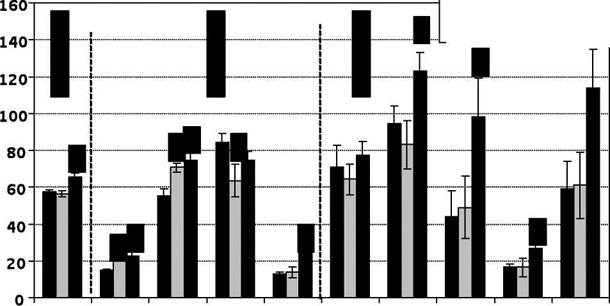
□ Атеросклероз ■ Таурин GABAAsp /lOCIu/100 Туг Тгр /100 Туг DA 5-HIAA*Trp 5-HT* Рис. 5 Концентрации нейроактивных аминокислот, биогенных аминов, их предшественников и производных в отделах мозга крыс на фоне экспериментального атеросклероза и введения таурина, 650 мг/кг (нмоль/г) Кроме этого, при введении Таи в среднем мозге и стриатуме (рис.5) наблюдался рост содержания триптофана, тенденция к повышению содержания 5-НТ и 5-HIAA, что свидетельствует об активирующем воздействии соединения на серотониновую систему исследованных структур центральной нервной системы при экспериментальном атеросклерозе, опосредованном повышением содержания предшественника — триптофана [105]. Введение Таи приводило также к нормализации в среднем мозге уровня тирозина, который был снижен при атеросклерозе (рис. 5). Так как известно, что повышением доступности тирозина удается активировать синтез катехоламинов в головном мозге [107,108], можно предполагать модулирующее влияние Таи на эти показатели, что подтверждается повышением содержания дофамина в стриатуме (рис.5). Практически важным выводом из представленных исследований следует считать наличие у Таи адаптогенного действия на процессы формирования фонда свободных аминокислот в и биогенных аминов при атеросклерозе. Тем самым очевидно, что механизм действия Таи при атеросклерозе не исчерпывается его изветными эффектами. Одновременно, учитывая сложность моделирования у крыс развёрнутых проявлений атеросклеротических поражений сосудов, нами для сравнения воспроизведена также модель экспериментального атеросклероза у кроликов [108], что позволило вызвать у них макроскопические изменения со стороны крупных артерий, приводящие к нарушениям регионарного кровотока, и в результате — метаболические нарушения, являющиеся следствием хронической гипоксии тканей, а не только самого атеросклероза как системной обменной патологии обмена веществ. Целесообразно при этом напомнить, что практически все известные клинические признаки атеросклероза проявляются лишь при формировании ишемических расстройств, возникновение которых возможно при наличии макроскопически определяемых поражений сосудов [109]. Макро- и микроскопическое исследование аорты кроликов опытной группы в данном эксперименте подтвердили воспроизведение её атеросклеротического поражения с выраженной картиной липоидоза [НО]. В описываемой ситуации экспериментального атеросклероза в плазме крови и тканях кроликов нами выявлен выраженный аминокислотный дисбаланс, характеризующийся снижением содержания предшественника Таи цистина, а также гликогенных аминокислот (серина, аспартата, глутамина, валина), лейцина и этаноламина. Полученные результаты свидетельствуют о дефиците серусодержащих аминокислот, не опосредованном снижением уровня метионина, их основного предшественника [37]. Снижение на этом фоне, как и при моделировании атеросклероза у крыс, соотношения АРУЦ и ароматических аминокислот (рис.6), также является признаком угнетения антитоксической функции печени [99]. Таким образом, характер аминокислотного дисбаланса при экспериментальном атеросклерозе у кроликов (рис.6) практически не отличается от такового у крыс (рис.4), у которых воспроизведены в основном метаболические нарушения, свойственные этой патологии.
Поиск по сайту: |


 Комплексный параметр для а-фазы, ч"1
Комплексный параметр для а-фазы, ч"1 После орального введения Таи он быстро всасывается с максимумом концентрации в крови к 10 минуте (рис.1), но средние значения радиоактивности, как и следует ожидать, при этом ниже, чем при внутривенном введении препарата. Однако, после достижения максимума при назначении Таи per os, его концентрация экспоненциально снижается примерно с такой же скоростью, как и при внутривенном введении. С учётом всасывания динамика концентраций 14С- Таи при его оральном назначении может быть описана уравнением 2:
После орального введения Таи он быстро всасывается с максимумом концентрации в крови к 10 минуте (рис.1), но средние значения радиоактивности, как и следует ожидать, при этом ниже, чем при внутривенном введении препарата. Однако, после достижения максимума при назначении Таи per os, его концентрация экспоненциально снижается примерно с такой же скоростью, как и при внутривенном введении. С учётом всасывания динамика концентраций 14С- Таи при его оральном назначении может быть описана уравнением 2:

 ia^aiii
ia^aiii
 Процентное содержание таурина от суммы свех свобождных аминокислот
Процентное содержание таурина от суммы свех свобождных аминокислот

 ■ Контроль
■ Контроль