|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Активація мРНК. Приєднання 43S комплексу до мРНК (48S-передініціаторний комплекс)
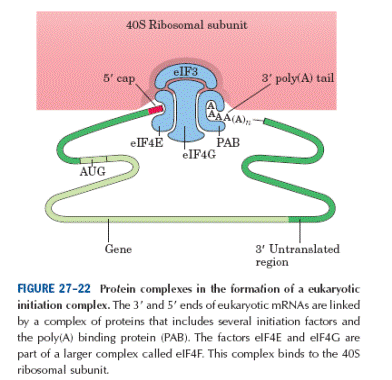
Як ми вже знаємо, мРНК приєднується до 43S комплексу. При цьому велике значення відіграє також фактор еІF3. Вважають, що еІF3 бере участь у формуванні центру, який розпізнає мРНК, з’єднуючись з певними факторами групи, що готують мРНК до трансляції, доведено- з фактором eIF -4G. Отже підготовка еукаріотичної мРНК до інціціації відбувається поетапно: комплексeIF -4F , а саме eIF- 4Езв’язується з кеп-структурою мРНК;eIF -4G координує роботу всіх факторів; eIF 4В формує комплекс за участі eIF- 4A, індукуючи її РНК-залежну геліказну активність- розплітання вторинної структури 5'-кінця мРНК з використанням енергії гідролізу АТФ. eIF- 4A існує у вільному і зв’язаному у комплексі eIF- 4Fстанах,є найчисленнішими факторами ініціації еукаріотів.
Перебіг подій під час зв’язування з сар-мРНК та кеп-ініційованого розплітання 5’-НТО (див. рис. вище): 1) 4Е субодиниця комплексу 4F з’єднується оборотно з сар-структурою мРНК (Кд=10-9 М), структурним скафолдом виступає 4G, який об’єднує роботу 4А і 4Е. 2) 4В формує хеліказний комплекс з eIF 4Fіактивує хеліказну активність 4А за участі АТФ (РНК-залежна АТФазна активність фактору 4А). 3) На наступному етапі- вивільнення 4Е, 4G, 4А після розщеплення АТФ. 4В залишається у комплексі. 4) Відбувається приєднання розчинної форми eIF 4А · АТФ і подальша хеліказна реакція. 5)
Отже, активована за участі факторів групи 4F та 4В, мРНК здатна приєднати рибосомний передініціаторний 43 S комплекс. Це відбувається за рахунок взаємодії 4G з еІF-3 та полі –А – зв´язуючими білками, а принципово направляється кеп-звязуючим білком 4Е.
Таким чином, за представленого перебігу подій утворюється 48S ініціаторний комплекс. Він містить 40S субодиницю, мРНК, потрійний комплекс- [метіоніл-тРНК, еІF-2· ГТФ], еІF-3, еІF-4В, еІF-1, еІF-1А, еІF-4F. Однак антикодон ще не зв’язаний з ініціаторним кодоном АУГ. 3. Сканування 5´-НТО в пошуках ініціаторного АУГ та його розпізнавання Асоціація рибосомних субодиниць з мРНК і розпізнавання ініціаторного кодону у еукаріотів розмежені в просторі і часі. В той же час в більшості випадків ініціаторний кодон може бути віддаленим від 5’-кінця на кілька десятків нуклеотидів. В еукаріотичній мРНК перший АUG триплет, який має відповідний контекст (GCC (A/G) CC AUG G), є ініціаторним (послідовність Marilyn Kozak). Бувають випадки, що не завжди перший від 5’- кінця триплет АUG є ініціаторним кодоном, оскільки знаходиться не в консенсусному оточенні Marilyn Kozak. Майже всі функціональні ініціаторні AUG кодони еукаріотичної мРНК мають перед собою триплет, що розпочинається з пуринового нуклеотидного залишку (A/G). Найчастіше це аденіловий нуклеотид. Отже при скануванні мРНК з 5’-кінця 40S субодиниця з ініціаторною аміноацил-тРНК розпізнає наведену вище структуру: 5´ GCC (A/G) CC AUG G 3´ Якщо замість A(G) стоїть С(U), то це -слабка ініціація. Рибосома проскакує і ініціації не відбуваються. З 3’-кінця від стартового кодону повинна йти G-послідовність. На основі ряду даних Marilyn Kozak запропонувала гіпотезу, згідно з якою 43S комплекс взаємодіє з кепованим 5’-кінцем мРНК (беруть активну участь білки комплексу 4F, а також на особливу роль заслуговуютьеІF-1,еІF-1А).еІF-1 єнеобхідним для розпізнавння контексту АУГ. Отже як при об'єднанні, так і при скануванні є необхідним повний комплект білкових факторів, а комплекс, що утворився (нагадаю), 48S ініціаторний комплекс. Далі 43S комплекс рухається по мРНК, залишаючи 4Е на кепі, читає її послідовність, поки не зустрінеться триплет AUG в контексті Marilyn Kozak. Цей триплет розпізнається ініціаторною аміноацил-тРНК. Тут комплекс зупиняється і проходить наступний етап ініціації, а саме приєднання 60S субодиниці до комплексу. Конформаційні зміни приводять до вивільнення ряду факторів ініціації (див. презент). Гіпотеза на сьогодні блискуче доведена. Різні теорії свідчать про існування двох моделей старту процесу ініціації. 1). Згідно з старою класичною моделлю - спершу до кеп-структури мРНК приєднується комплекс еІ- 4F- перші кроки підготовки 5'-НТО мРНК, далі процес подальшого розплітання… А 43S передініціаторний комплекс сідає на вже підготовлену ділянку по сусідству з стартовим кодоном. 2). Більш реальна за умов in vivo та загальноприйнята модельMarilyn Kozak - 4Е утримується кеп-структурою, в той час як 4А, 4В і 4G асоціюють з 43S передініціаторним комплексом. Таким чином саме 40S рибосомна субодиниця, що "несе" всі білкові фактори ініціації, сідає в районі кеп- кінця мРНК і сканує мРНК, стимулючи хеліказну і АТФазну активність білкових факторів, у пошуках стартового ініціаторного кодону.
Поиск по сайту: |

 Повторювання актів взаємодії мРНК- зв’язаного фактору 4В з 4А·АТФ сприяють розплітанню всіх прилеглих нижче структур мРНК. Саме ці цикли сприяють перебуванню 5’-НТО в стані одноланцюгової (без шпильок) структури.
Повторювання актів взаємодії мРНК- зв’язаного фактору 4В з 4А·АТФ сприяють розплітанню всіх прилеглих нижче структур мРНК. Саме ці цикли сприяють перебуванню 5’-НТО в стані одноланцюгової (без шпильок) структури.