|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Быстродействующие механизмы предвосхищения
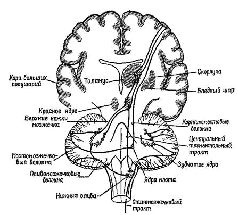
Признав факт существования произвольных движений, мы должны ответить на вопрос: как же они осуществляются? Ответ на этот вопрос можно дать на основе анализа процессов предвосхищения, с одной стороны, и раскрытия сущности предпрограммирующих процессов, подготавливающих результаты ряда движений прежде, чем они будут реализованы, — с другой. Вспомним, что в главе V указывалось, что для процессов предвосхищения необходим только приблизительный план, черновой набросок тех операций, которые должны быть выполнены. Как только будет выработан такой приблизительный план движений, дальнейшую их отшлифовку может взять на себя обычная сервосистема обратных связей. На основании нейро-анатомических и нейрофизиологических данных можно допустить, что некоторые аппараты нервной системы идеально приспособлены для обеспечения этой общей планирующей функции; и, как показывают результаты поведенческих экспериментов, эти функции действительно осуществляются такими аппаратами. Центральным звеном этой управляющей системы являются филогенетически новейшая часть мозжечка, эта удивительная структура, панорама которой открывается взору лишь при определенном сечении (см. Рис. XII—9, XII—10).
Как уже отмечалось выше, передняя часть мозжечка развилась при переходе морских позвоночных к наземному образу жизни. До этого момента мозжечок функционировал главным образом как дополнительный вестибулярный контролирующий механизм. Рыбы поддерживают ориентацию в соответствии с осью своего тела; их движения начинаются от этой основной линии. Амфибия и другие земноводные имеют дополнительную основную линию, это земля под ними, «низ», определяемый посредством гравитационных сил. Поэтому для организма становятся полезными новые вертикальные ориентиры позы и вследствие этого развивается передняя часть мозжечка. Наконец, приматы в известной мере освобождаются от гравитационной основной линии благодаря развитию двигательных навыков и приспособляются к древесному образу жизни и прямой позе. Теперь полушария мозжечка становятся доминирующими. У человека повреждение этих отделов мозга ведет к нарушению координации движений, к грубым промахам при слежении за целью и к атаксии при выполнении произвольных движений.
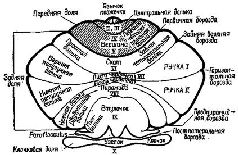
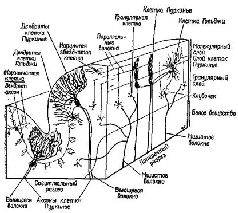
К счастью для нейрофизиологии, полушария мозжечка имеют удивительно гомогенное строение (см. Рис. XII—11) и поэтому относительно просты для анализа. Их основными связями являются: 1) связи через спинной мозг со структурами, обеспечивающими афферентацию движений; 2) связи с корой больших полушарий (см. Рис. ХII—9). Благодаря этим связям полушария мозжечка могут функционировать, как компаратор, сопоставляющий сигналы, поступающие от коры больших полушарий, с сигналами, идущими от периферии. Сущность этих процессов сравнения становится особенно интересной в свете последних детальных анатомических и физиологических данных (см. Рис. XII—12). Полушария мозжечка имеют кору, которая представляет собой как бы «прямоугольную решетку», напоминающую по своей конфигурации вертикальные колонки, обнаруженные в коре больших полушарий. Правда, в отличие от коры все эти мозжечковые решетчатые структуры фактически идентичны. Решетка построена таким образом, что, минуя один или два синапса, все поступающие в мозжечок импульсы преобразуются в тормозные сигналы. Этот невероятно развитый тормозной механизм должен служить следующим целям: «Это исключительное превращение всех поступающих сигналов в тормозные посредством самое большее двух синаптических реле придает мозжечку с точки зрения воздействия на входной сигнал характер аппарата, заглушающего колебания в системе. Вследствие этого становится невозможным непрерывное хранение информации посредством импульсов, циркулирующих по сложным нейрональным путям, как ото имеет место в коре больших полушарий или на путях между корой и базальными ганглиями. Через 30 мсек после поступления сигнала в кору мозжечка вызванные разряды уже не регистрируются. Остальные отделы центральной нервной системы требуют от мозжечка, по-видимому, не осуществления сложных реверберационных циклов в пределах самого мозжечка, а быстрой и четкой реакции на поступление какого-либо частного вида информации» (Eccles, Ito, Szentagothai, 1967, p. 311).
Быстрое стирание содержания записи предполагает, что последовательность операций сличения выполняется тоже довольно быстро. Эта особенность системы выходит за рамки обычного сервомеханизма обратных связей, в котором для длительного поддержания устойчивого состояния требуется постоянная информация о результатах движения. Т. Рач в своей работе, опубликованной в «Экспериментальной психологии» (1951), правильно сформулировал эту проблему, причем значительно раньше, чем были собраны соответствующие факты. «Процесс циркуляции возбуждения по замкнутому кругу от коры больших полушарий к (нео)-мозжечковым структурам выполняет функцию не столько устройства, корригирующего ошибки, сколько составляет часть механизма, который может мгновенно осуществлять предвосхищение влияния. Подобный замкнутый контур, хотя и не информированный о конечных результатах движения, мог бы, как говорится, «вчерне» программировать движение и таким образом сократить досадные издержки, связанные с коррекцией движения посредством аппарата обратных связей, который использует поступающую извне информацию» (Ruch, 1951, р. 205). Основная функция мозжечка, связанная с объемом регистрации, заключается в том, чтобы составить схему движения без детальной его разработки. В действительности такое кратковременное состояние, которое аналогично тому, что создается порой больших полушарий, формируется на фоне спонтанной активности. «Таким образом, следует предположить, что даже в условиях минимального сенсорного «входа» существует состояние динамического равновесия в уровне активности различных типов нейронов мозжечка. Отрицательный образ обобщенного результата работы коры мозжечка формируется в ходе процесса, аналогичного созданию скульптуры из камня. Форма, обладающая пространственно-временными признаками, достигается постепенно посредством набора тормозных импульсов, подавляющих «бесформенные» фоновые разряды нейронов, расположенных на более низких уровнях, подобно тому как вечные формы в скульптуре создаются путем очень избирательного удаления резцом всего лишнего из первоначально бесформенного камня» (Eccles, Ito, Szentogothai, 1967). Подробнее я остановлюсь на этом приборе и на его аналоге — в коре больших полушарий в главе XIII. Здесь же мы должны только констатировать, что механизмы предвосхищающего’ схематичного планирования движения можно рассматривать в качестве эквивалента быстродействующего компьютера, оценивающего конечный результат последовательной серии движений. Сходство между голограммой и процессом формирования образа заставляет предположить, что здесь имеет место процесс типа преобразования Фурье, однако и об этом речь будет идти в следующей главе. Во всяком случае, результаты предварительного расчета движений всегда немедленно поступают как к периферической мускулатуре, так и в моторную область коры больших полушарий через различные мозжечковые пути. С инженерной точки зрения подобный быстрый расчет ожидаемого «выхода» в настоящее время считается исключительно полезным приспособлением (Kelley, 1968). По-видимому, система «мозг-поведение» уже давно овладела им. Резюме Проблема мозговой организации поведения во многих отношениях является обратной стороной формирования Образа (восприятий и чувств). Вследствие повсеместного существования в нервной системе механизма обратной связи (и механизма предвосхищения) контроль над движениями достигается не непосредственно с помощью сигналов, адресующихся к сокращающимся мышечным волокнам, а посредством сигналов, адресующихся к рецепторам, которые регулируют мышечное сокращение. Таким образом, мышечные рецепторы становятся частью гармоничной системы (сервомеханизма Т—О—Т—Е), назначением которой является регуляция мышечных реакций в ответ яа изменения внешней среды. Короче говоря, нервная регуляция поведения осуществляется посредством воздействия на рещепторные процессы, а не через непосредственный контроль мышечных сокращений. Главная » Карл Прибрам. Языки мозга. Экспериментальные парадоксы и принципы нейропсихологии
Поиск по сайту: |