|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Активация как реакция на неопределенность
Что порождает эту неуравновешенность, это подавленное состояние духа? Факты говорят о том, что даже процесс зарождения таких состояний не прост. Переживания «интересов», мотивов (потребностей) и эмоций (аффектов) являются результатом состояний, возникающих, когда организм сталкивается с новизной — новизной, которая создается вследствие того, что непрерывно меняющийся «мир внутри нас» приходит в соприкосновение со всегда изменяющимся «миром вне нас». В результате исследований, проведенных в моей лаборатории с использованием техники разрушения отдельных участков мозга, удалось выделить по крайней мере два компонента ориентировочной реакции: один из них указывает на процесс исследования и опробования, а другой выявляется, когда организм фиксирует новизну. Только после такой фиксации возникает привыкание. Результаты этих исследований были получены примерно в следующей последовательности. Несколько лет назад мы показали, что изменения темперамента и личности, легко вызываемые удалением височных долей мозга, связаны с удалением амигдалы; эта структура височной доли, регулирующая потребности, обсуждалась в предыдущей главе (Pribram and Bagshaw, 1953; см. Рис. Х—3, стр. 215). Дальнейший анализ показал, что амигдала участвует в различных типах поведения, которые мы обозначили как четыре фактора, развивая идеи Кеннона о реакциях «агрессии и бегства», возникающих при электрической стимуляции участков гипоталамуса (Pribram, 19606). Наши четыре фактора, дополняющие классификации Кеннона, включают пищевое и половое поведение. Поскольку между амигдалой и гипоталамическими структурами существуют тесные анатомические связи, эти результаты казались вполне приемлемыми до тех пор, пока я не почувствовал недостаточность описательной корреляции между анатомией мозга и поведением и не попытался понять механизм этих отношений.
Если бы я удовлетворился тем, чтобы изучать поведение, как таковое, я, как и другие исследователи, поставил бы перед собой вопрос, не реализуется ли пищевое поведение, агрессия, бегство и половое поведение различными участками амигдалы. Когда были проведены эксперименты (с удалением или стимуляцией), направленные на разрешение этого вопроса, был получен, в сущности, отрицательный ответ. Но то, что я хотел узнать, касалось психологического процесса, того общего, что характеризовало эти виды аффективного поведения, поскольку одиночное разрушение (даже какой-нибудь целой анатомической формации) могло бы изменить одним ударом все эти различные типы поведения. Концепция «инстинкта» (Beach, 1955), несмотря на ее правдоподобность, по многим причинам была неудовлетворительной. Например, она не может объяснить, почему решающее влияние на изменение агрессивного поведения после удаления амигдалы оказывают социальные факторы (см. Рис. XI—2). Это могут объяснить различные формы концепции «влечения» вследствие наличия сильных связей между механизмами амигдалы и гипоталамуса. Однако, как отмечалось ранее, такие концепции «влечения» также не в состоянии объяснить влияние разрушения и стимуляции гипоталамуса. Поэтому я решил избрать противоположный подход к проблеме и поставить вопрос следующим образом: будет ли поведение, которое явно не является врожденным или управляемым влечениями, испытывать на себе влияние удаления амигдалы? Проведенные эксперименты не имели прямого отношения к исследованию упомянутых форм аффективного поведения. В сотрудничестве с Дж. Шварцбаумом (Schwartzbaum and Pribram, 1960), Мюриель Бэгшоу (Bagshaw and Pribram, 1965) и Э. Херстом (Hearst and Pribram, 1964a, 19646) я провел эксперименты по переносу навыка. В одной серии экспериментов обезьян обучали выбирать из двух интенсивностей серого цвета более светлый. Затем во второй серии экспериментов, в которой проверялся перенос навыка, более светлая панель пары предъявлялась в сочетании с еще более светлой. Взятые для контроля нормальные обезьяны в соответствии с выработанным у них навыком продолжали выбирать из двух панелей более светлую. Обезьяны же с удаленной амигдалой во время тестовых испытаний действовали случайным образом, как если бы они решали совсем новую задачу (см. Рис. XI—3). В другом эксперименте на перенос навыка обезьян ббучали выбирать из двух нарисованных квадратов квадрат большего размера, а затем проверяли, будут ли обезьяны выбирать большую из двух окружностей. Контрольные животные решали эту задачу, а животные с удаленной амигдалой — нет. Кроме того, в ситуации выработки оперантных условных реакций было проанализировано явление генерализации стимула. Обезьяны обучались регулировать освещение в экспериментальной камере, нажимая с разной частотой на педаль. Обезьяны с удалением амигдалы не отличались от контрольных и обнаружили такой же эффект генерализации, когда освещение в камере ступенчато менялось по сравнению с тем, на которое они первоначально были натренированы. (Это поразительное отсутствие связи между переносом навыка и явлением генерализации уже само по себе интересно, но к рассматриваемому вопросу не имеет отношения.)
При выборе этих задач мы исходили из предположения, что их решение не требует участия гипоталамуса. Поскольку в обоих экспериментах при удалении амигдалы перенос навыка нарушался, один из моих выводов состоял в том, что процесс, контролируемый амигдалой, отличен от тех, которые обычно связывают с функцией гипоталамуса. Ключом к разгадке того, что же представляет собой этот процесс, могут служить наблюдения за обезьянами, выполняющими задание с переменой стимулов. Как отмечалось ранее, обезьяны, лишенные амигдалы, не способны ни к переносу навыка, ни к выбору стимула по абсолютной величине. Во время тестовых экспериментов они вели себя, как в совершенно новой ситуации, и действовали на начальной стадии наугад (Douglas, 1966; Schwartzbaum and Pribram, 1960). Продолжая эти наблюдения (Bagshaw and Benzies, 1968; Bagshaw and Coppock, 1968; Bagshaw, Kimble and Pribram, 1965; Bagshaw and J. Pribram, 1968; Kimble, Bagshaw and Pribram, 1965), мы показали, что в действительности при удалении амигдалы менялись реакции обезьян на новизну. Как по поведенческим, так и по электроэнцефалографическим показателям привыкание было длительным. С другой стороны, разрушение амигдалы вело к устранению висцеральных показателей ориентировочного рефлекса (КГР, изменение в частоте сердцебиения и дыхания) при отсутствии повреждения самих механизмов, ответственных за возникновение висцеральных реакций (см. Рис. XI—4). Эти результаты говорят о том, что ориентировочный рефлекс на новизну проходит через две гипотетические стадии: первая, характеризующаяся поведенческими ориентировочными реакциями, «сравнивает», устанавливает новизну; вторая, связанная с висцеральными реакциями, ведет к «фиксации» в нервной системе нового раздражителя и тем самым создает основу для привыкания (Pribram, 1969). Таким образом, нарушения в поведении агрессии и бегства, пищевом и половом поведении, наблюдаемые в результате удаления амигдалы, я истолковал как результат тех затруднений, которые возникают в работе механизма, фиксирующего следы в нервной системе. Этот механизм необходим для организации поведения во времени. Проверка этой интерпретации предполагала, что поведение отсроченного чередования (выбор между двумя одинаковыми ящиками для пищи, которые заполнялись в последовательности справа — слева — справа — слева и так далее при последовательных пробах) (Pribram, Lim, Poppen and. Bagshaw, 1966) также окажется нарушенным, и это предсказание подтвердилось. Судя по этим результатам, амигдала выполняет гораздо большее число функций. Но это еще не все. Обращение к психологическому процессу фиксации помогло объяснить до сих пор не объяснимое наблюдение (Miller, Galanter and Pribram, 1960, см. Главу 14). В главе X говорилось о больной, у которой была произведена двусторонняя амигдалэктомия. За год после операции она сильно прибавила в весе. Казалось, что этот случай давал блестящую возможность выяснить непосредственно у больной, каковы были ее ощущения, заставлявшие ее есть так много. Она обычно отвечала, что ощущает очень немногое — она не чувствует, что очень голодна, то есть она не ощущала работу контролирующей и фиксирующей системы. Такая потеря регистрации — обычное явление в клинике у больных эпилепсией, возникающей вследствие повреждений вокруг амигдалы, повреждений, которые также порождают известные феномены deja vu (неадекватное переживание чего-то как уже знакомого) и jamais vu (неадекватное переживание известной ситуации как незнакомой). Таким образом, можно сказать, что контролирующие Образы характеризуют наши переживания, касающиеся «мира вне нас», во многом так же, как перцептивные образы характеризуют его свойства. Эти эксперименты и их анализ позволяют подойти к рассмотрению второй, более важной и широко распространенной точки зрения на то, каким образом возникают переживания мотивов и эмоций, и прежде всего те переживания интереса, побуждений и аффектов, которые зависят от механизма активации (arousal). Только что упомянутые результаты экспериментов ставят вопрос о том, при каких условиях активация (измеряемая с помощью той же самой техники и тех же критериев, которые были использованы при исследовании амигдалы) приводит к фиксации и привыканию и при каких — к их нарушению. Классический ответ, который был дан на этот вопрос (Lindsley, 1951; Hebb, 1955), состоит в том, что уровень активации определяет его конечный эффект. Сейчас к этому можно добавить, что в результате изучения ориентировочных реакций стало известно, что уровень активации зависит от организации, конфигурации ожиданий, от состояния мозга, с которыми сопоставляется новый сигнал. Таким образом, уровень активации следует понимать как степень соответствия и несоответствия между конфигурациями, степень организации или дезорганизации, а не как величину возбуждения, которое меняется само по себе (см. Hebb, 1949; Лурия, 1960). С тех пор как для характеристики активации начали привлекать понятие степени организации, стало возможным выразить уровень активации через понятия информации и неопределенности. Количество информации или неопределенности выражается числом ответов «да» — «нет», необходимым для ответа на вопрос, являются ли два объекта воспринимаемого или переживаемого поля одинаковыми или различными. Неопределенность — обратная сторона информации; термин «неопределенность» употребляется как противоположный термину «информация», которая прямо пропорциональна определенному числу бит (двоичных единиц «да» — «нет»). Чем больше это число, тем меньше неопределенность системы. Таким образом, можно было бы сказать, что такая точка зрения на теорию активации внесла в нее некоторую меру неопределенности.
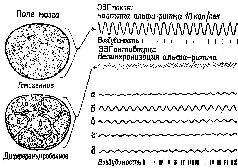
Само понятие «уровень» у сторонников теории активации, судя по их анализу, весьма неопределенно. Уровень активации — это не какое-то количественное изменение интенсивности, энергетического уровня центральной нервной системы, а нарушение равновесия, организации, измеряемой количеством неопределенности (и, следовательно, количеством информации), которое характеризует системы. На нейронном уровне активация сопровождается изменением динамических структур возбуждения в ретикулярной формации ствола мозга (см. Huttenlocher, 1961) и в коре (Burns, 1968, см. Главу 5), а не общего количества спайковых разрядов нейронов. Даже первоначальное определение активации как электроэнцефалографической реакции основано на том наблюдении, что в ЭЭГ возникает низковольтная высокочастотная активность и что такая активность указывает на десинхронизацию. Как было отмечено Линдсли (1961), это предполагает, что во время десинхронизации нервные элементы становятся функционально независимыми друг от друга и, таким образом, способными функционировать в качестве отдельных каналов, проводящих различную информацию. Эту возросшую сепарацию можно описать как увеличение степени организации или в терминах теории информации как увеличение неопределенности (см. Рис. XI—5). Естественным следствием такого понимания было бы предположение, что активация представляет собой состояние, при котором независимая активность нейронных ансамблей создает свободу для «резонанса» с другими, которые, несмотря на пространственную удаленность, имеют с ними общие характеристики. Таким образом, рассредоточенная информация в состоянии активации легче направляется по нужному адресу. Гипотеза о том, что активация является в какой-то мере механизмом неопределенности, может показаться нам привлекательной. Но это еще не все, что известно в отношении активации. Например, в эксперименте, направленном на изучение того, какое влияние в ранний период жизни оказывает ограничение движений мы обнаружили, что инактивация младенца сопровождается увеличением лабильности автономной нервной системы (Konrad and Bagshaw, 1970). Но прежде чем будут выяснены отношения между неопределенностью и контролем над поведением, нам необходимо еще раз вернуться к теории сервомеханизмов. Результаты Шехтера еще не получили своего объяснения: усиления того или другого переживания в результате инъекции адреналина можно достигать путем количественного изменения состояния неопределенности, вызываемого введением определенного агента; специфичность же таких чувств, как враждебность или дружелюбие и так далее, еще требует объяснения.
Поиск по сайту: |