|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Витаминные коферменты II группы
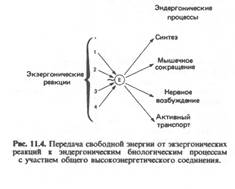
1.Тиаминдифосфат (ТДФ или тиаминпирофосфат). Кофермент состоит из двух гетероциклов (ядра тиазола и пиримидинового цикла) и пирофосфатного остатка. Тиаминдифосфат в составе ферментов катализирует окислительное декарбоксилирование пировиноградной и альфа-кетоглутаровой и перенос гликольальдегидной группы от кетосахаров на альдосахара в составе фермента транскетолазы. 2. Коэнзим А (КоА-SH), кофермент ацилирования(кофермент ацетил- и ацилтрансфераз)-участвует в метаболизме углеводов ( декарбоксилирование пирувата и альфа-кетоглутарата), в реакциях окисления и синтеза жирных кислот, холестерина, гема, ацетилхолина, обезвреживании чужеродных веществ и др. КоА-SH - это типичный нуклеотид и состоит из аденилового нуклеотида соединенного с пантотеновой кислотой (витамин В3)и бета-меркаптоэтиламином: Функция КоА- SH состоит в активации и переносе остатков карбоновых кислот СН3 - СООН + АТФ + КоА-SH → СН3- СО∼S-КоА + Н2О Уксусная кислота активная уксусная кислота (ацетил-КоА) 3.Пиридоксальфосфат (ПАЛФ) и пиридоксаминфосфат (ПАМФ)являются производным витамина В6. ПАЛФ принимает участие в реакциях обмена аминокислот, переносит химические группы (NH2, CO2): 4.Биоцитинсостоит из биотина (витамина В8) и остатка лизина. Биотин это гетероцикл состоящий из имидазольного и тиофенового колец. Биоцитин является коферментом ферментов карбоксилаз, которые присоединяют СО2 к молекуле карбоновой кислоты с удлинением цепи на 1 атом углерода. 1.Принимает участие в реакциях карбоксилирования с участием АТФ: например, образование малонил-КоА – интермедиата в синтезе жирных кислот или образование щавелевоуксусной кислоты, метаболита цикла трикарбоновых кислот. 2.Биоцитин принимает участие в реакциях транскарбоксилирования (без участия АТФ), при которых субстраты обмениваются карбоксильными группами: R1 - COOH + R2H R1H + R2 - COOH 5.Тетрагидрофолиевая кислота (ТГФК).Структура кофермента включает кольцо птеридина, пара-аминобензойную кислоту (ПАБК) и глутаминовую кислоту. ТГФК синтезируется из витамина В9 (фолиевой кислоты) под действием фермента дигидрофолатредуктазы, который присоединяет 4 атома водорода к фолиевой кислоте. Этот фермент ингибируется противоопухолевым препаратом метотрексатом, что ведет к торможению синтеза нуклеотидов, необходимых для репликации ДНК и тормозит рост опухоли. Бактерии в отличие от животных способны самостоятельно синтезировать фолат из предшественников – птеринового ядра, ПАБК и глутамата. Сульфаниламидные антимикробные препараты структурно подобны ПАБК и поэтому тормозят синтез ФК, задерживая размножение бактерий. Функции ТГФК состоят в межмолекулярном переносе одноуглеродных фрагментов: 1. Метильного (-СН3 ), ТГФК является синергистом другого кофермента – метилкобаламина, который также переносит этот важный остаток. 2. Метиленового ( - СН2 - ) 3. Метинильного ( - СН = ) 4. Оксиметильного ( - СН – ОН ) 5. Формильного ( - СОН ) 6. Форминоиминного ( - СН=NH ) ТГФК принимает участие в обмене аминокислот (синтез метионина из гомоцистеина), в синтезе дезоксирибонуклеотидов (тимидилата для ДНК путем метилирования дУТФ) и пуриновых ядер (аденина и гуанина), холина, креатина, адреналина. Макроэргические связи Вообще термин «макроэргическая связь» используется исключительно для связей, энергия которых используется в метаболизме и не указывает на истинную величину энергии связей. Обычно это относится к макроэргическим молекулам — биологическим молекулам, которые способны накапливать и передавать энергию в ходе реакции. При гидролизе одной из связей высвобождается более 20 кДж/моль. По химическому строению макроэрги — чаще всего ангидриды карбоновой и фосфорной кислот, а также других слабых кислот. Примеры макроэргических соединений — молекулы АТФ. Связь между тремя остатками фосфорной кислоты. При разрыве этих связей выделяется очень много энергии - 40кДж при отрыве каждой молекулы. В этом и заключается энергоемкость АТФ. Как известно, энергия связей всегда положительна, другими словами, разрыв любой связи, в том числе и макроэргической, всегда требует дополнительных затрат энергии, но их результирующая - выше. Важно отметить, что последовательное усложнение материи неизбежно приводит к усложнению и усовершенствованию инструментов её самоуправления и управления. При этом количество таких инструментов весьма ограничено физическими и химическими свойствами самой материи. Появление макроэргических связей на горизонте эволюционного процесса, открыло фактически новую эру в возможностях мобильного применения химических (биохимических) связей в формировании, поддержании и усовершенствовании гомеостатического устройства в инструментальном обеспечении жизнедеятельности организованной материи. Одним из таких структурно-функциональных инструментов является межмолекулярное взаимодействие. Понятие «макроэргическая связь» учитывает энергетический эффект преобразованной связи посредством химической реакции вещества с нормальными свойствами. Связи между остатками фосфорной кислоты являются макроэргическими – при их гидролизе выделяется энергия. Такие связи принято обозначать волнистой черточкой. Энергия 1-й молекулы АТФ может служить только для 1-й реакции. АДФ и АМФ – не способны быть источником энергии. Разрыв макроэргических связей в молекулах сопровождается выделением энергии, используемой для биосинтеза и транспорта веществ, мышечного сокращения, пищеварения и других процессов жизнедеятельности организма. Макроэргические соединения (macroergic compounds) АТФ: Химическая энергия гидролиза АТФ может использоваться для сопряжения с эндоэргическими процессами.
В АТФ цепочка из трех фосфатных остатков связана с 5'-OH-группой аденозина. Фосфатные группы обозначаются как α, β и γ. Рибоза связана с α-фосфатом фосфоэфирной связью. Три фосфатных остатка соединены между собой менее устойчивыми фосфоангидридными связями. При физиологических значениях рН АТФ несет четыре отрицательных заряда. Собственно действующим коферментом является комплекс АТФ с ионом Mg2+, координационно связанным с α- и β-фосфатом (Mg2+ ּ АТФ4-, на рисунке не показан). Для простоты чаще всего говорят только об АТФ.
Изменение свободной энергии ΔGo' гидролиза фосфоангидридных связей в АТФ при рН 7 в стандартных условиях составляет от -30 до -35 кДж/моль.
Роль АТФ: Все молекулы АТФ в клетке непрерывно участвуют в каких-либо реакциях, постоянно расщепляются до АДФ и вновь регенерируют. ü Биосинтез веществ ü Транспорт веществ через мембрану ü Изменение формы клетки и её движение Эти процессы вкупе с процессом образования АТФ получили название АТФ-цикл: кругооборот АТФ в жизни клетки. Нуклеозидфосфаты, родственные АТФ И АДФ. Нуклеозидфосфаты (1) являются не только исходными соединениями в биосинтезе нуклеиновых кислот, они обладают также функциями коферментов, служат для запасания энергиии участвуют в цепи переноса энергии в эндоэргических процессах. Метаболические интермедиаты часто становятся реакционноспособными («активированными») при присоединении фосфат содержащих остатков (фосфорилирование). Так, присоединение нуклеозиддифосфатных остатков делает реакционноспособными исходные соединения в синтезе полисахаридов и липидов . Лигазы катализируют сшивание соединений за счет энергии нуклеозидтрифосфатов.
Олиго-и полинуклеотиды полинуклеотид иначе нуклеиновая кислота (англ. polynucleotide) —полимерная молекула, состоящая из нуклеотидов. Описание Полинуклеотиды — биополимеры, нуклеиновые кислоты (от лат. nucleus — ядро), образованные нуклеотидными звеньями, которые в свою очередь состоят из азотистого основания, углеводного остатка и фосфатной группы. Цепочки из нуклеотидов соединяются через остаток фосфорной кислоты (фосфодиэфирная связь). Полинуклеотиды, составленные из рибонуклеотидных звеньев, называютсярибонуклеиновыми кислотами (РНК), из дезоксирибонуклеотидных мономеров —дезоксирибонуклеиновыми кислотами (ДНК). Различаются они не только по типу углеводного остатка, но и по типу азотистых оснований. В состав всех нуклеиновых кислот входят аденин, цитозин и гуанин, а также тимин (ДНК) и урацил (РНК). При обозначении полинуклеотидов указывают сокращенные названия нуклеотидных звеньев в направлении 5' → 3', т. е. слева направо. Если фосфатная группа одного нуклеотида взаимодействует с 3'-OH-группой другого нуклеотида, образуется динуклеотид с фосфодиэфирной связью. Такой динуклеотид несет на 5'-конце свободную фосфатную группу, а на 3'-конце — свободную OH-группу. Поэтому за счет образования еще одной фосфодиэфирной связи можно присоединить новый мононуклеотид. Таким путем образуются олигонуклеотиды и, наконец, полинуклеотиды. В клетках синтез нуклеиновых кислот осуществляется ферментами, которые образуют новые цепи полинуклеотидов, используя в качестве матрицы резидентную молекулу нуклеиновой кислоты. Полинуклеотиды определенного нуклеотидного состава могут быть использованы как структурные элементы ДНК-наноструктур или элементы биогенных наночастиц. Иллюстрации
Защитный, эффект при введении загадочных спиралей может сохраняться от нескольких дней до (в отдельных случаях) недель. Самым поразительным оказалось, что противовирусный эффект полинуклеотидов в большинстве случаев много выше, чем это можно было бы ожидать, судя по количеству обнаруживаемого интерферона. Если мы введем в экспериментальную систему то количестве защитного белка интерферона, которое стимулирует в ней применение синтетических полинуклеотидов, то уровень защиты от введенного интерферона будет намного ниже. В чем тут дело, толком никому еще не ясно. Возможно, что противовирусное действие полинуклеотидов много сложнее, чем просто стимуляция интерферона.
Между тем спектр действия интерфероногенов также необычайно широк, как и самого интерферона. И не однажды у исследователей возникала мысль: не могут ли они оказаться полезными в борьбе против вирусов, вызывающих образование опухолей? Такие эксперименты на животных проводились в некоторых лабораториях. Сообщали о явном профилактическом действии интерфероногенов при. заражении животных опухолевыми вирусами. Иногда удавалось предотвращать образование опухолей и даже вызывать их рассасывание. Следует все же отметить, что опыты такого рода ни в коей мере не вышли еще за рамки лабораторных экспериментов. Изучение интерферона и интерфероногенов еще далеко от завершения. Но все чаще мы получаем добрые вести с этого участка фронта борьбй е вирусными инфекциями.
Олигонуклеотиды-полимер короткий нуклеиновых кислот, как правило, с 20 или меньше базы. Хотя они могут быть созданы путем расщепления облигаций более сегментов, они являются в настоящее время более часто синтезирован полимеризации отдельных нуклеотидов прекурсоров. Автоматизированная синтезаторы позволяют синтез олигонуклеотидов до 160 до 200 базы. Длина олигонуклеотидных обычно обозначается «mer» (от греческого «МЕРОС», «часть»). Например фрагмент 25 баз будет называться 25-МЭ. Потому что олигонуклеотиды легко связать их соответствующих дополнительных нуклеотидов, они часто используются как датчики для обнаружения ДНК или РНК. Примеры процедур, использующих олигонуклеотиды ДНК microarrays, южной части пятна, Асо анализ, флуоресцентные in situ гибридизация (рыба) и синтез искусственных генов. Олигонуклеотиды, состоящий из ДНК (oligodeoxyribonucleotides) часто используются в цепной реакции полимеразы, процедура, которая может значительно усилить практически любой небольшой фрагмент ДНК. Там олигонуклеотидных именуется как грунт, позволяя ДНК-полимеразы для расширения олигонуклеотидных и воспроизвести дополнительные пряди. Антисмысловые олигонуклеотиды являются одной нити ДНК или РНК, которые являются дополнительными к выбранной последовательности. По Антисмысловые РНК они препятствуют белков перевод некоторых матричной РНК нити путем привязки к ним. Антисмысловые ДНК может использоваться для конкретной дополнительной (кодирования или не кодирование) РНК. Если привязка места Этот гибрид ДНК и РНК может снизиться, фермент RNase х. Синтез олигонуклеотидов — это химический синтез относительно коротких фрагментов нуклеиновых кислот с заданной химической структурой (последовательностью). Метод очень полезен в современной лабораторной практике, поскольку он позволяет получать олигонуклеотиды нужной последовательности быстрым и недорогим способом. В то время какферменты синтезируют ДНК и РНК в направлении 5'→3', химический синтез олигонуклеотидов проводится в обратном направлении (3'→5'). В настоящее время процесс реализован как твердофазный синтез по амидофосфитному методу с использованием амидофосфитных строительных блоков, производных защищённых 2'-дезоксинуклеозидов (dA, dC, dG и T),рибонуклеозидов (A, C, G и U) или химически модифицированных нуклеозидов, например, LNA. Для получения целевого олигонуклеотида строительные блоки последовательно конденсируют с растущей олигонуклеотидной цепью в порядке, задаваемом последовательностью олигонуклеотида. Процесс был полностью автоматизирован в конце 1970-х гг. После завершения синтеза цепи продукт отделяется от твердофазного носителя и защитные группы удаляются, после чего олигонуклеотид очищается ВЭЖХ. Протекание побочных реакций накладывает ограничение на длину синтезируемого олигонуклеотида (до 200 нуклеотидных остатков), поскольку число ошибок накапливается с увеличением длины целевого продукта. Одноцепочечные молекулы ДНК или РНК длиной 15-25 оснований являются наиболее востребованными синтетическими олигонуклеотидами. Олигонуклеотиды находят широкое применение в молекулярной биологии и медицине, например, как антисмысловые олигонуклеотиды, праймеры для секвенирования и амплификации ДНК, зонды для определения комплементарных последовательностей ДНК и РНК, инструменты для нацеленного введения мутаций и сайтов рестрикции, а также для синтеза искусственных генов. В процессе эволюции синтеза олигонуклеотидов было разработано четыре основных метода создания связей между нуклеозидами. Детальное рассмотрение этих методов сделано в подробных литературных обзорах
Поиск по сайту: |


 Структура АТФ:
Структура АТФ: 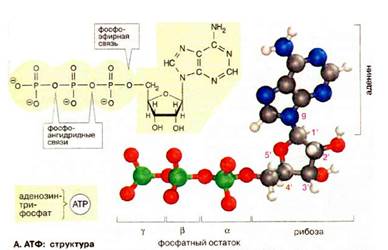

 Свободная энергия гидролиза высокоэнергетических связей.
Свободная энергия гидролиза высокоэнергетических связей.



 На рис. Показано, как через эти различные нуклеозид- и дезоксинуклеозидтрифосфаты энергия и строительные блоки передаются на определенные метаболические пути, где они используются для биосинтеза липидов, белков и в первую очередь для биосинтеза ДНК и РНК.
На рис. Показано, как через эти различные нуклеозид- и дезоксинуклеозидтрифосфаты энергия и строительные блоки передаются на определенные метаболические пути, где они используются для биосинтеза липидов, белков и в первую очередь для биосинтеза ДНК и РНК.
 Нуклеотидные звенья соединяются через фосфатную группу двумя сложноэфирными связями: с 3'-атомом углерода одного нуклеотида и с 5'-атомом - другого.
Поликонденсация множества нуклеотидов приводит к образованию полинуклеотидов (нуклеиновых кислот). Главная цепь всех полинуклеотидов построена по единой схеме – она содержит остатки моносахарида (сахара) и фосфорной кислоты. Поэтому главную цепь макромолекулы называют сахарофосфатной.
Нуклеотидные звенья соединяются через фосфатную группу двумя сложноэфирными связями: с 3'-атомом углерода одного нуклеотида и с 5'-атомом - другого.
Поликонденсация множества нуклеотидов приводит к образованию полинуклеотидов (нуклеиновых кислот). Главная цепь всех полинуклеотидов построена по единой схеме – она содержит остатки моносахарида (сахара) и фосфорной кислоты. Поэтому главную цепь макромолекулы называют сахарофосфатной.
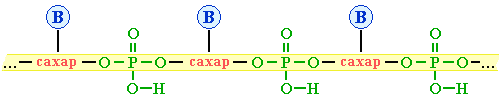
 – Азотистые (пуриновые и пиримидиновые) основания: аденин (А), гуанин (Г), тимин (Т), цитозин (Ц), урацил (У), которые связаны с главной цепью как заместители. Схематически структуру макромолекул нуклеиновых кислот можно представить формулой:
– Азотистые (пуриновые и пиримидиновые) основания: аденин (А), гуанин (Г), тимин (Т), цитозин (Ц), урацил (У), которые связаны с главной цепью как заместители. Схематически структуру макромолекул нуклеиновых кислот можно представить формулой:
 При этом учитывается, что подобные структурные звенья отличаются входящими в их состав азотистыми основаниями (В = А, Г, Т, Ц, У).
Полинуклеотиды относят к кислотам, т.к. в каждом структурном звене их макромолекул содержится остаток ортофосфорной кислоты, определяющий кислотные свойства за счет диссоциации связи О-Н. Последовательность нуклеотидных звеньев в полинуклеотидной цепи характеризует первичную структуру нуклеиновых кислот, которую записывают в виде однобуквенных обозначений азотистых оснований (или соответствующих нуклеотидов):
... – А – Г – Ц – Т – Ц – А – Т – ...
При этом учитывается, что подобные структурные звенья отличаются входящими в их состав азотистыми основаниями (В = А, Г, Т, Ц, У).
Полинуклеотиды относят к кислотам, т.к. в каждом структурном звене их макромолекул содержится остаток ортофосфорной кислоты, определяющий кислотные свойства за счет диссоциации связи О-Н. Последовательность нуклеотидных звеньев в полинуклеотидной цепи характеризует первичную структуру нуклеиновых кислот, которую записывают в виде однобуквенных обозначений азотистых оснований (или соответствующих нуклеотидов):
... – А – Г – Ц – Т – Ц – А – Т – ...
 Решение этой задачи — дело будущего. Пока же большинство исследователей полагают, что в основе профилактического, а также обнаруживаемого иногда и лечебного действия этих препаратов лежит интерферон. Хотя достоверно установлено, что они могут, помимо всего прочего, и другими путями влиять на состояние противовирусной защиты организма человека, например, активизировать его иммунные системы, способствовать выработке защитных противовирусных антител.
Решение этой задачи — дело будущего. Пока же большинство исследователей полагают, что в основе профилактического, а также обнаруживаемого иногда и лечебного действия этих препаратов лежит интерферон. Хотя достоверно установлено, что они могут, помимо всего прочего, и другими путями влиять на состояние противовирусной защиты организма человека, например, активизировать его иммунные системы, способствовать выработке защитных противовирусных антител.