|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Сравнительная физиология дыхания животных ⇐ ПредыдущаяСтр 2 из 2
Физиологическое значение дыхания для живых организмов Дыхание является неотъемлемой функцией любых живых организмов, поскольку оно обеспечивает поступление в организм кислорода и выведение углекислого газа. Кислород необходим для осуществления окислительных процессов, в ходе которых высвобождается энергия разрушенных химических связей, часть которой аккумулируется в макроэргических связях АТФ и в дальнейшем может использоваться клетками для выполнения различных видов работы (синтеза собственных веществ, транспорта ионов и некоторых молекул против концентрационных градиентов, механической работы и т.д.). Очень немногие организмы (некоторые простейшие, бактерии, водоросли, грибы, фораминиферы, паразитические круглые черви) способны обеспечивать себя энергией, высвобождающейся в результате бескислородного расщепления глюкозы и поэтому способны выживать в бескислородных условиях. Однако эффективность бескислородного расщепления глюкозы ничтожно мала и поэтому такой способ образования энергии возможен только у низкоорганизованных очень мелких организмов с очень низким уровнем метаболизма. У большинства же животных для нормальной жизнедеятельности необходимо поступление кислорода из окружающей среды, которое осуществляется в процессе дыхания. Кроме того, в результате расщепления органических веществ до неорганических образуется СО2, Н2О и аммиак (при расщеплении белков и аминокислот). Образующаяся в результате катаболизма вода частично включается в водный баланс организма, частично выводится через специальные органы выделения; азотсодержащие неорганические вещества также выводятся специализированными органами выделения; СО2 в больших количествах является весьма токсичным для организма и в связи с этим должен постоянно выводиться из организма благодаря дыханию.
молочнокислые бактерии
лямблия фораминиферы
аскарида гриб Fusarium poae (культура)
Рис. 1. Примеры анаэробных живых существ
Таким образом, дыхание является одним из ключевых процессов жизнедеятельности любых живых организмов (в том числе и растений, который, наряду с фотосинтезом, тоже дышат), поскольку обеспечивает поступление в организм из окружающей среды кислорода, постоянно расходующегося в ходе окислительных процессов, и выделение углекислого газа, постоянно образующегося в ходе катаболизма.
Основные способы и органы дыхания у животных Осуществление дыхания у разных животных во многом зависит от их уровня организации, размера, особенностей внешних покровов, способа жизни, интенсивности метаболизма и многих других обстоятельств. Так, у очень мелких организмов (наибольший размер тела которых не превышает 1 мм) дыхание возможно путем диффузии газов непосредственно через всю поверхность тела и последующую диффузию газов внутри самого тела (т.е. такие животные не требуют наличия специальной транспортной системы). При этом в связи с очень малыми размерами, несмотря на низкую скорость диффузии газов, она будет эффективной для поступления кислорода вглубь организма и выделения углекислого газа наружу.
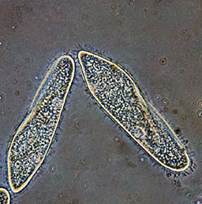
амеба обыкновенная инфузория-туфелька эвглена зеленая Рис. 1. Пример очень мелких организмов, у которых дыхание осуществляется непосредственно путем диффузии газов через поверхность тела Вместе с тем, если размеры животного близки к 1 см и форма тела близка к сферической, диффузия газов через поверхность их тела внутрь окажется неэффективной для нормального обеспечения кислородом глубоких слоев тела. В связи с этим для таких животных уже необходима не просто диффузия газов с поверхности тела через внутреннее его содержимое, а конвекционный перенос газов внутри организма (т.е. необходима специальная система транспорта газов в организме). Единственным исключением из этой закономерности являются некоторые относительно крупные беспозвоночные с большой относительной поверхностью тела, но при этом очень малой толщиной, а также беспозвоночные с очень низким уровнем метаболизма. Так, медузы, несмотря на свои относительно большие размеры и близкую к сферической форму тела, дышат через поверхность тела и не имеют специальной транспортной системы. При этом медузы характеризуются весьма низкой интенсивностью обмена веществ, в связи с чем количества кислорода, поступающего путем диффузии через поверхность тела вглубь к тканям, достаточно для сохранения жизнедеятельности. Некоторые медузы, хотя и могут иметь весьма крупные размеры, но содержат всего 1% (от общей массы тела) органического вещества (остальное – соли и вода) и характеризуются очень низким уровнем метаболизма; к тому же метаболически активные их клетки расположены очень близко к поверхности тела, что делает возможным эффективное поступление кислорода к ним путем простой диффузии через поверхность тела. Наряду с медузами, способностью получать кислород через поверхность тела и доставлять его вглубь путем диффузии обладают некоторые относительно крупные кишечнополостные (гидры, губки, коралловые полипы) и плоские черви, живущие в воде. Дыхание путем диффузии газов у этих животных при отсутствии специальной транспортной системы возможно благодаря относительно большой удельной площади поверхности их тела, но при этом очень малой его толщины. В частности, у этих беспозвоночных, как правило, очень удлиненное, но при этом уплощенное (почти нитевидное) тело или имеется множество разнообразных плоских выростов, увеличивающих площадь поверхности тела. Благодаря весьма тонкому телу возможна эффективная диффузия кислорода на относительно короткие расстояния вглубь тканей.
медуза гидра губка
коралловый полип плоский червь Рис. 2. Примеры животных, которые, несмотря на относительно большую площадь поверхности тела, способны дышать путем диффузии газов через поверхность тела и последующей диффузии газов внутри самого организма Таким образом, поступление кислорода через поверхность тела (а не через специальные дыхательные поверхности) и последующая его доставка к тканям путем диффузии по градиенту парциального напряжения (а не благодаря конвекционному переносу кровью) возможны только либо для ничтожно мелких беспозвоночных, либо для беспозвоночных с очень низким уровнем метаболизма. Характер поступления кислорода в организм также во многом зависит от размеров тела, характера их покровов и способа жизни. В частности, для некоторых животных с влажной хорошо проницаемой для газов и воды кожей (некоторые амфибии, черви) возможно дыхание непосредственно через поверхность тела без участия специальных органов дыхания. При этом у плоских червей в связи с очень узким (не более 1 см в диаметре) и длинным телом дальнейшее поступление кислорода к внутренним тканям возможно путем диффузии через собственные ткани, тогда как у амфибий в связи с более крупными размерами для доставки газов к клеткам требуется специальная транспортная система.
безлегочные саламандры Рис. 3. Примеры животных, способных во влажной среде дышать через поверхность тела, но при этом для доставки кислорода к тканям использующих конвекционный перенос газов жидкостями
Большинство животных (те, что характеризуются большими размерами тела или более высоким уровнем метаболизма, чем, у тех, что дышат путем диффузии газов через поверхность) нуждаются в специальных органах дыхания – специализированных поверхностях для газообмена. Эти специальные поверхности для газообмена, как правило, более тонкие, чем остальные покровы тела, и гораздо лучше кровоснабжаемые. Они поставляют кислород в кровеносное русло или непосредственно к тканям (трахейная система у насекомых начинается от дыхалец, сообщающихся с окружающей средой, и доставляет кислород путем конвекционных потоков воздуха в ней непосредственно к клеткам тканей). Организация органов дыхания у животных во многом зависит от среды их обитания. Так у большинства первичноводных животных дыхание может осуществляться как через поверхность тела (у относительно просто организованных беспозвоночных), так и с участием жабр (органов, специализирующихся на дыхании в воде, они омываются водой и густо кровоснабжаются). У наземных животных для дыхания может служить кожа (у относительно немногих), легкие (у большинства наземных позвоночных) и система трахей (у насекомых). Если специализированная дыхательная поверхность, являющаяся производным поверхности тела, вывернута наружу, образуя выпячивание, такой орган дыхания называют жаброй. Вторично жабры могут быть заключены в специальные полости (оперкулярные полости), как это имеет место у рыб. Если же специализированная дыхательная поверхность (производное поверхности тела) образует углубление (или впячивание), направленное внутрь тела, его называют легким. В самом простом случае легкие представляют собой простые мешковидные углубления, в которых осуществляется газообмен (как например, у легочной наземной улитки). В более сложном случае они имеют чрезвычайно сложную разветвленную структуру, по своей площади во много раз превышающую площадь поверхности тела животного (легкие позвоночных). У насекомых дыхательная система представлена системой разветвленных трубочек (трахей), берущих начало от специальных отверстий (дыхалец) на поверхности тела и подходящих ко всем частям тела. Таким образом, у насекомых дыхательный орган представляет собой комбинацию распределительной системы (системы трахей) с газообменной системой (самые тонкие трахеи, через стенки которых возможна диффузия газов непосредственно к клеткам тела).
Легкие позвоночных
Легкие амфибий и рептилий
Легкое птицы
Схема строения легкого млекопитающего
Трахейная система насекомых
участие кожи в дыхании у амфибий Рис. 4. Разнообразные органы дыхания у животных В целом, жабры чаще служат для дыхания в воде, а легкие – для дыхания в воздухе. Вместе с тем, есть и исключения из этой закономерности: например, голотурии имеют водные легкие, в которых осуществляется основной газообмен. Жабры могут быть модифицированы для дыхания в воздухе, хотя у большинства рыб при извлекании их из воды жабры вследствие малой механической жесткости и прочности спадаются и слипаются под действием собственного веса, в связи с чем поверхность, открытая для воздуха, оказывается во много раз меньше той, что доступна действию воды, что сильно препятствует поглощению кислорода и приводит к удушью рыб, несмотря на то, что содержание кислорода в воздухе гораздо выше такового в воде.
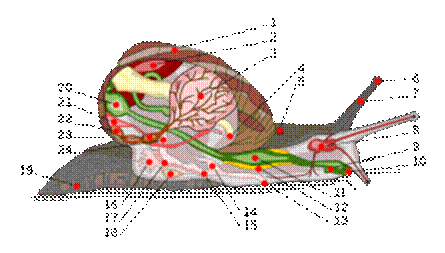
Дыхание животных в воде с помощью жабр Строение жабр у разных первичноводных животных Жабры – специализированные органы дыхания у большинства первичноводных животных, представляющие собой относительно тонкие и густо кровоснабжаемые выпячивания определенных участков поверхности тела, у рыб погруженные в специальную оперкулярную полость. В самом простом случае жабры представляют собой выпячивание наружу тонкого участка поверхности тела, густо снабженного капиллярами. Такие примитивные жабры имеются у кольчатых червей на параподиях (у свободно-подвижных форм) или на головном конце тела (у кольчатых червей, живущих в трубках). У большинства высших ракообразных жабры расположены на верхних отделах грудных ног или на боковых стенках тела.
жаброногий рачок
кольчатый червь
Рис. 5. Фото беспозвоночных, имеющих примитивные жабры на своих конечностях
У водных личинок насекомых имеются т.н. трахейные жабры — тонкостенные выросты на различных частях тела, в которые заходит густая сеть слепо заканчивающихся трахей.
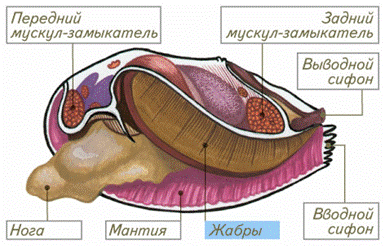
У большинства моллюсков жабры – ктенидии, представляющие собой парные двусторонне перистые жабры, расположенные в мантийной полости. При этом каждый ктенидий состоит из опорного тяжа и двух рядов ресничных пластинок (ламмелл), придающих ктенидию сходство с птичьим пером. Внутри опорного тяжа залегают приносящий и выносящий кровеносные сосуды, мышцы и нервы ктенидия. Собственно газообмен с водой происходит через эпителий ламелл. У головоногих моллюсков в основании каждого ктенидия находится дополнительное жаберное сердце, сокращения которого усиливают кровоток.
Рис. 6. Схема строения моллюска (видны жабры, представляющие собой видоизмененные участки мантии)
Из иглокожих (исключительно морских донных животных, большей частью свободноживущих, реже сидячих, встречающихся на любых глубинах Мирового океана) жабры имеются у морских ежей и морских звёзд. Причем жабры иглокожих представляют собой выросты кожи, в которые заходит полость тела (это т.н. кожные жабры, выполняющие дыхательную функцию). Их стенка очень тонкая, поэтому через неё легко происходит газообмен.
Рис. 7. Фото иглокожих, имеющих кожные жабры
У голотурий (морских огурцов или кубышек) формируются особые органы дыхания — водные лёгкие, представляющие собой пару ветвистых мешков, открывающихся в кишечник около анального отверстия. Через это отверстие осуществляется пропускание воды (закачивание и выкачивание) в кишечник, необходимое для газообмена. Эти животные при стрессе часто выбрасывают через анальное отверстие заднюю часть кишки вместе с водными легкими (эвисцерация); затем утраченные структуры регенерируют.
Рис. 8. Фото иглокожего (голотурии), имеющей водные легкие
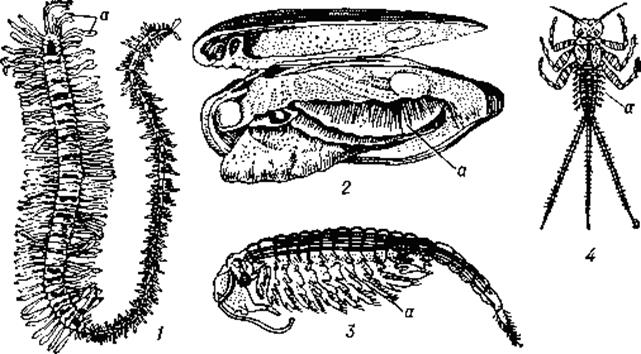
Рис. 9. Дышащие жабрами беспозвоночные: 1 — многощетинковый червь; 2 — моллюск перловица с приоткрытой раковиной и частично удалённой складкой мантии; 3 — жаброногий рачок бранхипус; 4 — личинка подёнки; а — жабры.
У всех первичноводных хордовых животных в глотке имеются ряды парных отверстий, т.н. жаберные щели, открывающиеся наружу непосредственно или через околожаберную полость. Так, у кишечнодышащих (морских животных типа полухордовых, длина тела которых составляет от нескольких см до 1 м; тело червеобразное, состоит из 3 отделов: хоботка, воротника и туловища, у основания хоботка, с брюшной стороны расположено ротовое отверстие, которое ведёт в глотку) боковые стенки пищевода пронизаны жаберными щелями (отсюда название кишечнодышащие), число которых достигает 200. Вода проникает через ротовое отверстие в пищевод и вступает в газообмен с кровью сосудов, густо пронизывающих перегородки между жаберными щелями. Аналогичным образом устроены органы дыхания у бесчерепных и оболочников. Так, у ланцетника (бесчерепные) жаберные щели (около 100 штук), пронизывающие глотку, открываются не прямо наружу, а в специальную околожаберную полость. У асцидии (оболочники) жаберные щели из глотки ведут в особую полость, выстланную энтодермой и состоящую из двух половин, сросшихся на брюшной стороне с мантией. Эта полость называется околожаберной, атриальной или перибранхиальной. Она залегает с каждой стороны между глоткой и наружной стенкой тела. Часть ее образует клоаку. Эта полость не является полостью тела животного. Она развивается из особых впячиваний наружной поверхности внутрь тела. Околожаберная полость сообщается с внешней средой при помощи клоакального сифона.
баланоглосс (кишечнодышащие)
ланцетник (бесчерепные), его жаберные щели, пронизывающие глотку, открываются не прямо наружу, а в специальную околожаберную полость
Асцидии (оболочники)
Рис. 10. Фото некоторых хордовых, имеющих жаберные щели, пронизывающие определенные отделы пищеварительной трубки
У круглоротых (миксины, миноги) в стенке глотки имеются жаберные отверстия, ведущие в жаберные мешки, стенки которых несут тонкие жаберные лепестки. В жаберных лепестках расположены кровеносные сосуды, в которых происходит газообмен. У миног каждый жаберный мешок открывается наружу жаберным отверстием, а у миксин жаберные отверстия соединены с продольным каналом, который открывается одной парой пор на боковых поверхностях тела.
Миноги миксина
Рис. 11. Фото круглоротых, жабры которых связаны с пищеварительной трубкой
У рыб жабры имеют более сложное строение, обеспечивающее значительное увеличение площади их респираторной поверхности, а также противоток воды и крови в жаберных пластинках. Кроме того, жабры рыб заключены в жаберную полость, обеспечивающую защиту этих относительно крупных органов и позволяющую обтекать жаберную поверхность наиболее эффективным образом.
Площадь дыхательной поверхности жабр у рыб зависит от уровня их метаболизма, во многом определяемого двигательной активностью. Так, у быстро плавающей макрели жаберная поверхность, отнесенная к единице массы тела, в 50 раз больше, чем у медлительной придонной рыбы морской черт.
макрель
морской черт
Рис. 12. Внешний вид макрели (быстро плавающая пелагическая рыба) и морского черта (медленно плавающая придонная рыба)
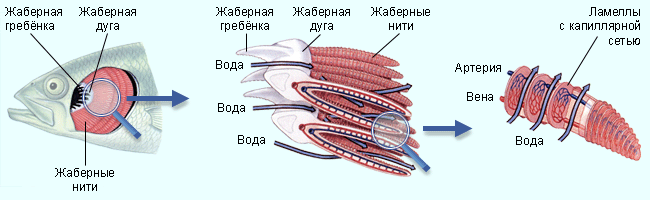
Жаберные щели рыб разделены жаберными дугами (как правило, имеется несколько жаберных дуг с каждой стороны). От каждой жаберной дуги отходят два ряда жаберных нитей (или жаберных перегородок). Кончики нитей, принадлежащих смежным жаберным дугам, соприкасаются, заставляя воду течь между нитями. На каждой жаберной нити располагаются ряды плотно уложенных жаберных пластинок (или лепестков). Газообмен между водой и кровью осуществляется в жаберных пластинках (дискообразной формы) по мере того, как вода протекает между ними в одном направлении, а кровь внутри жаберных пластинок – в другом. Жаберные пластинки свисают в жаберную полость. Жаберная полость снаружи прикрыта кожной складкой, укреплённой костями,— жаберной крышкой.
Рис. 13. Жабры рыб состоят из нескольких жаберных дуг. От каждой жаберной дуги отходит по два ряда жаберных нитей. На каждой нити имеются тонкие параллельные дискообразные жаберные пластинки. Внутри этих пластинок кровь течет в направлении, обратном току воды между пластинками (Randall, 1968).
Таким образом, респираторная поверхность жабр рыб значительно увеличена за счёт тончайших складок на поверхности жаберных пластинок. У личинок рыб и земноводных имеются наружные жабры, не имеющие жаберных крышек, а у личинок бесхвостых земноводных развиваются, кроме того, и своеобразные внутренние жабры.
Рис. 14. Жабродышащие позвоночные животные: 1 — асцидия (схема; с левой стороны частично удалены стенки тела, околожаберной полости и глотки): а — ротовой сифон, б — клоакальный сифон, в — жаберные щели; 2 — ланцетник (схематический продольный разрез): а — жаберные щели, б — околожаберная полость, в — кишка, г — наружное отверстие околожаберной полости; 3 — минога (продольный разрез через переднюю часть тела): а — жаберный мешок, б — кишка, в — рот, г — отверстие жаберного мешка в дыхательную трубку, д — межжаберные перегородки, е — сердце; 4 — акула (голова, вскрытая с брюшной стороны): а — жабры, б — жаберные щели, в — жаберные артерии, г — сердце; 5 — окунь (голова с удалённой жаберной крышкой): а — жабры, б — жаберные тычинки, в — сердце; 6 — личинка двоякодышащей рыбы лепидосирена: а — наружные жабры; 7 — личинка гребенчатого тритона (Triturus cristatus): a — наружные жабры; 8 — головастик лягушки Rana temporaria: a — наружные жабры, б — область внутренних жабр; 9 — хвостатое земноводное сирен: а — наружные жабры.
Характер тока воды и крови в жабрах рыб и его физиологическое значение Поток воды около жабры и поток крови внутри этого органа направлены противоположно друг другу (т.е. создается противоток воды, омывающей жаберную поверхность, и крови, протекающей в жаберных пластинках). Принцип противотока имеет важное значение для эффективной экстракции кислорода из омывающей жабру воды. Так, когда кровь уже готова покинуть жаберную пластинку, она встречает воду, только что вошедшую между жаберными пластинками (т.е. воду, из которой кислород еще не извлекался). Таким образом, эта кровь поглощает кислород из воды, в которой содержание кислорода еще не понизилось и в результате насыщение крови кислородом может достигнуть максимального уровня. По мере дальнейшего прохождения между пластинками вода встречает кровь со все более и более низким содержанием кислорода, и поэтому она будет все время отдавать кислород. Благодаря отмеченному противотоку в жабрах жаберные пластинки извлекают кислород из воды по всей длине, и вода, пройдя жабру, может потерять до 80-90% исходного количества кислорода (парциальное напряжение кислорода в крови, покидающей жабру, выше такового воды, покидающей пространство между жаберными пластинками). Это очень высокая степень экстракции кислорода (для сравнения в легких млекопитающих из альвеолярного воздуха экстрагируется около 25% кислорода). Если бы вода и кровь текли в одном направлении, такая высокая степень экстракции кислорода была бы невозможна, поскольку парциальное напряжение кислорода в крови могло бы в лучшем случае достичь того же уровня, что и в вытекающей из жаберной полости воды. Поскольку прокачивание воды через жабры требует энергии, благодаря увеличению экстракции кислорода противоток снижает энергетические затраты на прокачивание.
Наряду с рыбами противоток встречается и в жабрах крабов, но у них эффективность извлечения кислорода из воды намного меньше, чем у рыб. Например, у европейского берегового краба экстракция кислорода из воды, несмотря на наличие противотока, не превышает 7-23%. Низкая экстракция кислорода из воды в жабрах крабов отчасти может быть связана с более высоким жаберно-кровяным барьером для диффузии у них.
В связи с очень тесным расположением у рыб жаберных пластинок друг относительно друга (промежутки между ними могут не превышать 0,02 мм) первоначально возникла мысль, что для тока воды между жаберными пластинками создается очень большое сопротивление, что резко ограничивает ток воды. Вместе с тем, детальное изучение скорости тока воды в жабрах большинства рыб показало, что жаберные пластинки не создают высокого сопротивления. Так, поток воды, рассчитанный согласно уравнению Пуазейля, через жабры 150-граммового линя (Tinca) при давлении 5 мм водного столба должен быть равен 10,1 мл/с. Нормальный объем воды, прогоняемый через жабры этой рыбы, составляет 1-2 мл/с. Из этого очевидно, что жаберные пластинки не создают большого сопротивления, поскольку могли бы пропустить и больше воды (согласно теоретическим расчетам), чем реально проходит у живой рыбы. При этом наибольшие величины для потока воды найдены у некоторых антарктических рыб, очень необычных в том плане, что в их крови отсутствует гемоглобин. Сравнительно малое расстояние между жаберными пластинками (до 0,02 мм), в то же время не исключающее возможность прохождения воды между ними, обеспечивает очень тесный контакт жаберных пластинок и воды, что очень важно для эффективной экстракции кислорода кровью из воды.
Механизмы, обеспечивающие поддержание тока воды в оперкулярных полостях Для того чтобы жабры эффективно извлекали из омывающей их воды кислород, необходимы: ü постоянная смена воды у дыхательных поверхностей жабр (поскольку в отсутствии движения воды парциальное напряжение кислорода в ней у дыхательной поверхности жабры быстро снизиться вплоть до нуля и дальнейший газообмен будет фактически не эффективен) ü тесный контакт между водой и жаброй.
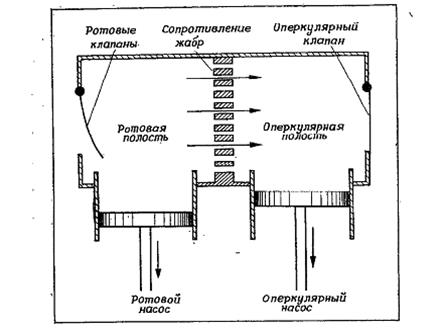
Для создания тока воды над поверхностью жабр существуют различные механические приспособления, способные обеспечивать движение воды 2-мя путями: ü движением жабры в воде ü перемещением воды вокруг жабры. Движение жабры в воде «практично» только для мелких организмов (например, для водных личинок некоторых насекомых, в частности, личинок поденок). Трудность движения жабры в воде у более крупных водных животных обусловлена большими затратами энергии при вращении жабры вокруг ее основания вследствие большого сопротивления воды; кроме того, несмотря на большие затраты энергии на движение жабры в воде, необходимо увеличивать и ее механическую прочность. Вместе с тем, некоторые относительно крупные водные животные используют этот способ для постоянной смены воды над дыхательными поверхностями жабр. В частности, крупная водная амфибия Necturus все же перемещает свои жабры, но эти движения происходят очень медленно. Движение воды над дыхательной поверхностью жабры – намного легче реализуемое решение. Такое движение может создаваться работой ресничек, как у простейших или в жабрах мидий и других двустворчатых моллюсков. Губки прогоняют воду через свои отверстия с помощью жгутиков. Вместе с тем, более распространены механические приспособления, подобные насосу, типичные для рыб и крабов. Так, костистые рыбы для прокачивания воды используют комбинированное действие ротовой полости и жаберных крышек при участии соответствующих клапанов для контроля потока (модель такого насоса представлена на рис.).
Рис. 15. Схема работы ротового и оперкулярного насосов, обеспечивающих непрерывное прокачивание воды через жабры. Воду через жабры рыб прогоняет двойная система насосов. При помощи соответствующих клапанов эти насосы обеспечивают однонаправленный ток воды около жаберной поверхности (Hughes, 1960).
Эта система для прокачивания воды через жабры состоит из 2-х насосов: ротовой полости и оперкулярной полости. Увеличение объема первого насоса (ротовой полости) достигается путем опускания челюсти, а увеличение объема оперкулярной полости (расположены по одной с каждой стороны, сообщаются с ротовой полостью) – за счет движения жаберных крышек при одновременном блокировании обратного тока воды вблизи краев кожными щитками, играющими роль пассивных клапанов. Действие этих 2-х насосов таково, что ток воды через жабры поддерживается на протяжении почти всего дыхательного цикла. Вода продолжает течь, несмотря на то, что во время части цикла давление в ротовой полости может сделаться меньше, чем во внешней среде, но при этом давление в оперкулярной полости поддерживается на еще более низком уровне, чем во рту. Таким образом, давление в ротовой полости на протяжении почти всего дыхательного цикла выше такового в оперкулярных полостях, что обеспечивает практически непрерывный ток воды через жабры. Вместе с тем, некоторые рыбы не способны дышать таким способом. В частности, крупные тунцы не выживают в неволе, если у них нет возможности беспрерывно плавать (в связи с этим их приходится помещать в большие кольцеобразные бассейны, где они могут плавать по кругу без остановки). Такие рыбы плавают, приоткрыв рот. При этом видимые дыхательные движения у них отсутствуют (т.е. ротовой и оперкулярный насосы не работают), но в связи с непрерывным плаванием с открытым ртом вода все равно непрерывно течет через жабры (это т.н. таранная вентиляция). Таранная вентиляция встречается не только у крупных, быстро плавающих пелагических рыб. Многие рыбы при малых скоростях используют для дыхания прокачивание, а при увеличении скорости переходят к таранной вентиляции. При этом переход к таранной вентиляции наблюдается в диапазоне скоростей от 0,5 до 1 м/с: при больших скоростях активные дыхательные движения прекращаются. Переход к таранной вентиляции не означает, что дыхание «пускается на самотек»; это значит только, что вентиляцию обеспечивают теперь не мышцы оперкулярного насоса, а плавательные мышцы тела и хвоста. При этом открытый рот увеличивает лобовое сопротивление, которое должно быть компенсировано усиленной работой мускулатуры. Однако возможно, что при высокой интенсивности дыхания, необходимой во время быстрого плавания, непрерывный ток воды при таранной вентиляции энергетически более выгоден, чем оперкулярное прокачивание. Степень открытия рта, по-видимому, регулируется таким образом, чтобы обеспечивать достаточный ток воды через жабры, но не увеличивать лобовое сопротивление больше, чем это требуется для адекватной вентиляции. К такому выводы пришли на основании экспериментов, в ходе которых макрелей заставляли двигаться с одной и той же скоростью, но постепенно снижали содержание кислорода в воде. При этом уменьшение содержания кислорода в воде приводило к постепенному раскрыванию рта, таким образом, что снижение содержания кислорода в воде компенсировалось увеличением тока воды через жабры. Если при протекании воды через жабры в них застревают твердые взвешенные частицы, то рыбы удаляют их за счет изменения направления тока воды. В частности, они расширяют ротовую полость (путем опускания нижней челюсти) при сомкнутых губах, что приводит к резкому снижению давления во рту и, как следствие, движению воды из оперкулярных полостей в ротовую полость (т.е. в обратном обычному направлении). Данный механизм аналогичен кашлю у млекопитающих, служащему для удаления слизи и твердых частиц из нижних дыхательных путей. Крабы прочищают свои жабры аналогичным способом. В частности, у крабов ток воды через жабры направлен почти всегда в одном направлении, но время от времени прокачивание прекращается и происходит внезапная реверсия потока за несколько секунд. Частота таких реверсий сильно варьирует: от каждой минуты до 1 раза в 10 минут или даже реже. По-видимому, эти внезапные реверсии потока служат для удаления частиц, застрявших в жабрах.
У некоторых животных движению воды около жабр способствует перемещение всего тела. Такой способ типичен для многих пелагических рыб. Так, у крупных тунцов жаберные крышки почти неподвижны, и необходимая высокая скорость потока воды над жабрами достигается путем быстрого плавания в воде. В связи с этим тунцы не могут выжить, если их лишить возможности плыть вперед, и поэтому при искусственном содержании их помещают в большие круглые бассейны, чтобы они могли беспрерывно двигаться, не встречая препятствий. У кальмаров и осьминогов тоже имеется тесная корреляция между локомоцией и током воды над жабрами. Вместе с тем, движение воды над жаберной поверхностью у рыб и кальмаров обеспечивается разными механизмами. Так, кальмар или осьминог вентилирует жабры, набирая воду в мантийную полость, а когда он выбрасывает ее через сифон, то продвигается в воде реактивным способом. В этом случае вентиляционная система была видоизменена для локомоции, но, как и у рыб, у кальмаров и осьминогов по мере того как с увеличением скорости плавания возрастает потребность в кислороде, последний автоматически поступает в большем количестве.
Другие функции жабр Наряду с участием в дыхании, жабры могут выполнять и другие функции. Так, анальные жабры у личинок комаров осуществляют осмотическую регуляцию (они всасывают из окружающей воды ионы, и эта функция является для них основной, тогда как участия в дыхании они не принимают), у рыб и крабов жабры также участвуют в осморегуляции (хотя основная их функция – дыхание), у двустворчатых моллюсков жабры, наряду с дыханием, участвуют в питании (они отцеживают как сито взвешенные в воде частички пищи, которые затем поступают в ротовую полость).
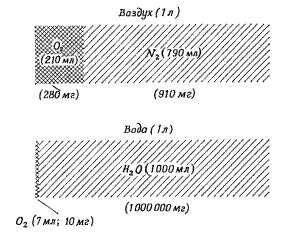
Дыхание наземных животных в воздухе Условия дыхания животных на суше Успешная адаптация животных в процессе эволюции к воздушному дыханию осуществилась в широком масштабе только у членистоногих и позвоночных. Кроме того, хорошо приспособлены к жизни на суше некоторые улитки (они встречаются даже в пустынях) и сравнительно небольшое число других беспозвоночных. Дыхание животных в воздушной среде имеет ряд преимуществ по сравнению с таковым в воде. Во-первых, воздух отличается от воды гораздо более высоким содержанием кислорода и соответственно большей его доступностью для живых организмов. Так, 1 л воздуха содержит 210 мл О2, который весит 280 мг. Оставшаяся часть воздуха – 790 мл N2 – весит 910 мг. Таким образом, для того, чтобы экстрагировать из воздуха, пропускаемого через органы дыхания, нужное количество кислорода, необходимо пропустить в 3,5 раза большую часть воздуха, чем вес необходимого кислорода. 1 литр пресной воды, находящейся в равновесии с атмосферным воздухом, при температуре 15°С содержит всего 7 мл кислорода. Эти 7 мл О2 весят 0,01 г (10 мг), что в 100 000 раз меньше веса воды. Таким образом, чтобы получить заданное количество кислорода, необходимо пропустить через дыхательные органы животного массу воды в 100 000 раз большую массы необходимого кислорода. Морская вода из-за присутствия солей характеризуется меньшей растворимостью газов, в связи с чем содержит еще меньше кислорода по сравнению с пресной (5,8 мл в 1 литре воды при температуре 15°С). При повышении температуры воды растворимость газов в ней уменьшается, что обуславливает снижение содержания газов в ней. Так, при 30°С пресная вода содержит 5,6 мл/л, а морская – 4,5 мл/л кислорода.
Рис. Схема, отражающая соотношение массы кислорода с массой атмосферного воздуха и воды. В воздухе содержится много кислорода (21%), а биологически инертный газ азот содержится в количестве, в 3,5 раза большем массы кислорода. В отличие от этого в воде растворенный кислород составляет всего 0,7% по объему (при 15°С и Ро2 в атмосферном воздухе 159 мм рт. ст.), что в 100 000 раз меньше веса инертной среды (воды).
Во-вторых, еще одно преимущество дыхания в воздухе – это сравнительно низкая его вязкость по сравнению с вязкостью воды. Так, вязкость воды при 20°С равна 1 сПз (сантипаузу), а вязкость воздуха – 0,02 сПз (т.е. вязкость воды в 50 раз больше вязкости воздуха). Более низкая вязкость воздуха по сравнению с водой облегчает передвижение воздушных масс и делает возможным перемену направления движения воздуха в воздухоносных путях и органах дыхания в процессе дыхательного цикла. Вместе с тем, у некоторых позвоночных, дышащих воздухом (птиц и насекомых), встречается однонаправленное движение воздуха в органах дыхания. В водных дыхательных системах движение воды почти всегда однонаправленное, поскольку если бы поток воды входил и выходил в одном месте или двигался то туда, то сюда, то большую часть воды приходилось бы ускорять, затем останавливать, затем снова ускорять, но уже в обратном направлении. Для беспрерывных изменений кинетической энергии воды требовались бы большие затраты энергии, поэтому у всех водных животных, получающих кислород из воды, движение воды в органах дыхания осуществляется в одном направлении. В-третьих, скорость диффузии кислорода в воздушной среде примерно в 10 000 раз больше, чем в воде при том же парциальном давлении (напряжении) кислорода. При такой быстрой диффузии размеры элементов дыхательного органа могут быть совсем иными, чем у животных, дышащих в воде. Так, расстояния, на которые газ диффундирует в легких, могут составлять несколько миллиметров, а диффузионные расстояния в жабрах рыб – ничтожные доли миллиметра. Несмотря на существующие преимущества дыхания животных в воздухе по сравнению с водной средой, существует и определенная трудность, связанная с испарением воды с поверхности воздухоносных путей и легких при дыхании. Вместе с тем, направленная внутрь сила для поглощения кислорода (159 мм рт. ст.) в 6 раз больше, чем сила для отдачи воды в сухой воздух (при 25°С – 25 мм рт.ст.). Даже при высоких температурах (около 37°С) давление водяных паров, обуславливающее отдачу воды в окружающую среду, мало по сравнению с давлением, заставляющим кислород поступать внутрь. Для того, чтобы не происходило чрезмерного испарения воды с дыхательной поверхности, наружный воздух не должен иметь к ней свободного доступа. В связи с этим газообменные поверхности обычно располагаются в специализированных дыхательных полостях (легких), что значительно ограничивает доступ воздуха. При этом обновление воздуха в такой полости часто очень тонко регулируется и не бывает более интенсивным, чем потребность их организма в кислороде. В связи с потерями воды вследствие испарения у наземных животных газообмен через всю поверхность тела встречается очень редко и, как правило, только во влажных местообитаниях. Например, дождевые черви, у которых весь дыхательный газообмен осуществляется через поверхность тела, очень чувствительны к недостатку воды. Они живут во влажных местах, и если остаются неприкрытыми на поверхности земли, то быстро высыхают и гибнут.
Лучшая доступность кислорода в воздухе по сравнению с таковой в воде обусловила более высокий уровень метаболизма у них и в связи с этим более высокий уровень структурной и физиологической организации. Кроме того, гораздо более низкая теплопроводность воздуха по сравнению с теплопроводностью воды сделала возможным формирование в процессе эволюции гомойотермии (в связи с лучшими условиями по поддержанию на постоянном уровне температуры тела). В то же время некоторые очень крупные и быстро плавающие рыбы способны поддерживать температуру своего тела на более высоком уровне по сравнению с температурой воды.
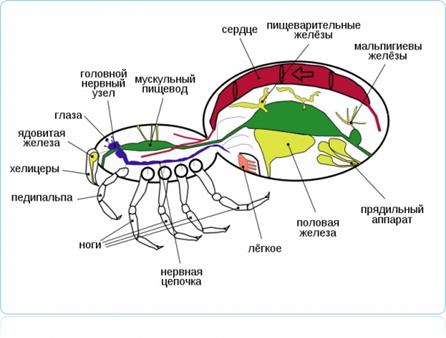
Органы дыхания у наземных животных У сухопутных животных встречаются 3 основных типа органов дыхания: ü легкие ü трахеи ü жабры. Жабры. Жабры довольно плохо приспособлены для дыхания в воздушной среде и используются лишь немногими животными – главным образом теми, которые сравнительно недавно перешли к сухопутному образу жизни и принесли с собой пережитки своего прежнего способа водного дыхания. Примером таких животных служат некоторые сухопутные крабы. Так, краб пальмовый вор (Birgus latro), почти полностью перешедший к наземной жизни (включая лазанье по кокосовым пальмам), имеет достаточно жесткие жабры, пригодные и для дыхания в воздухе. Другой сухопутный краб Cardiosoma может неопределенно долго жить и на суше и в воде, тогда как пальмовый вор через некоторое время погибает, если его держать погруженным в воду.
Краб пальмовый вор (Birgus latro)
мокрица Рис. Фото некоторых сухопутных членистоногих, способных дышать жабрами на суше.
Еще одна группа воздуходышащих ракообразных с жабрами – это наземные равноногие (Isopoda), называемые мокрицами. Эти животные предпочитают жить во влажных местах, и у видов, наиболее успешно приспособившихся к наземным местообитаниям, жабры заключены в полости, так что в функциональном плане их можно рассматривать как легкие. У некоторых рыб, способных дышать воздухом (но не у всех), функционирующие жабры сохранились. Так, обыкновенный угорь (Anguilla vulgaris) хорошо выживает на воздухе, если ему обеспечить достаточное охлаждение и влажность. Тогда большая часть кислорода поступает через кожу и меньшая – через жабры, поскольку жаберные нити частично слипаются, и в жаберной полости лишь небольшая часть их поверхности остается в контакте с воздухом. В результате угорь не получает обычного количества кислорода, и потребление кислорода уменьшается примерно вдвое.
Рис. Фото угря обыкновенного, который, несмотря на наличие жабр, приспособленных к дыханию в воде, способен выживать на воздухе благодаря жаберному и кожному дыханию.
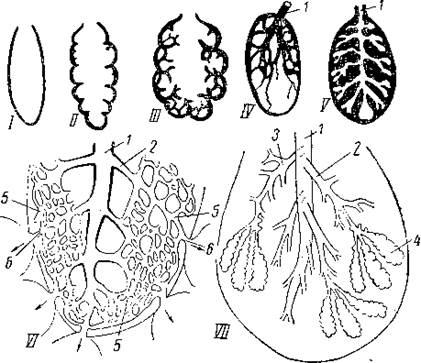
Легкие. Различают 2 типа легких: диффузионные и вентиляционные. Диффузионные легкие характеризуются тем, что обмен воздухом с окружающей средой осуществляется только путем диффузии. Легкие этого типа встречаются у относительно небольших животных, таких, как легочные улитки, скорпионы и некоторые равноногие раки. Диффузионные легкие у легочных улиток представляют собой часть мантийной полости, заполненную воздухом и имеющую густую капиллярную сеть. Такое легкое сообщается с окружающим воздухом через посредство единственного отверстия – пневмостома.
Рис. Схема строения легочной улитки. 1 — раковины; 2 — пищеварительная железа; 3 — лёгкое; 4 — анус; 5 — пневмостом; 6 — глаз; 7 — щупальце; 8 — мозг; 9 — радула; 10 — рот; 11 — зоб; 12 — слюнная железа; 13 — гонопор; 14 — пенис; 15 — влагалище; 16 — слизистая железа; 17 — яйцепровод; 18 — мешок любовных стрел; 19 — нога; 20 — желудок; 21 — почка; 22 — мантия; 23 — сердце; 24 — семявыносящий проток.
Легкие скорпиона имеют вид мешков, представляющих собой углубления в теле паука, закрывающиеся особыми отверстиями-дыхальцами. Одна стенка у легочных мешков похожа на книжку: состоит из множества тончайших «складочек-страничек». В эти складочки затекает гемолимфа и через тоненькую стенку поглощает из воздуха легочных мешков кислород и отдает углекислый газ.
Рис. Схема строения скорпиона, имеющего легкое. Вентиляционные легкие имеются только у позвоночных. Значительное и регулярное обновление воздуха в легких необходимо при больших размерах тела, сочетающихся с высокой интенсивностью метаболизма. Дыхательные системы позвоночных вентилируются входящими и выходящими (приливно-отливными) потоками воздуха. У птиц эти системы намного сложнее, чем у млекопитающих, и устроены таким образом, что и при вдохе, и при выдохе воздух может течь через легкие в одном направлении.
Вентиляционные легкие позвоночных
Трахеи. Этот тип органов дыхания характерен для насекомых. Он представляет собой систему трубочек, которые доставляют кислород непосредственно к тканям, так что циркуляция крови для транспорта газов оказывается ненужной. Газообмен в трахейной системе может происходить только путем диффузии, но у многих крупных и особенно высокоактивных насекомых имеет место и однонаправленное прокачивание воздуха через части трахейной системы. Преимущество однонаправленного потока состоит в том, что он обеспечивает намного лучший газообмен, чем при поочередном всасывании и выталкивании воздуха.
Роль кожи в дыхании наземных животных Газообмен через кожу является нормой и играет важную роль у амфибий, кожа которых влажная и имеет густую капиллярную сеть. У некоторых амфибий отсутствуют легкие, и газообмен у них осуществляется только через кожу. Так, некоторые мелкие саламандры (Plethodontidae) совсем не имеют легких, и весь газообмен осуществляется у них через кожную поверхность и отчасти через слизистую оболочку рта. Эти саламандры отнюдь не редкость – они составляют около 70% всех существующих видов саламандр. Это очень мелкие формы, встречающиеся как в наземных, так и в водных местообитаниях. Так, Desmognathus fuscus весит от 5 до 7 г, дышит в основном кожей, хотя около 15% всего газообмена осуществляется через слизистую оболочку рта и глотки. Кровь не отличается какими-либо особыми свойствами: и содержание гемоглобина, и его сродство к кислороду находятся в том же диапазоне, что и у других саламандр, как водных, так и наземных. У них нет специальных сосудов, которые несли бы насыщенную кислородом кровь от кожи к сердцу, и оксигенированная кровь, покидая кожу, смешивается с общей венозной кровью. У других амфибий в сердце имеется частичное разделение оксигенированной и венозной крови, но у безлегочных саламандр такого разделения, по-видимому, нет, и поэтому кровь у них никогда не бывает полностью насыщена кислородом. Тем не менее, широкое распространение безлегочных саламандр указывает на то, что они отлично адаптированы, несмотря на дыхательный аппарат, который, по всей видимости, является весьма несовершенным.
Рис. Фото мелкой безлегочной саламандры Typhlotpiton srelaeus.
При этом относительное значение кожи и легких в обеспечении организма кислородом у амфибий меняется на протяжении года. Так, зимой потребление кислорода очень невелико (в связи с низким уровнем метаболизма), и удельная роль кожи в поглощении кислорода выше, чем легких. Летом, когда в связи с повышением температуры и соответственно метаболизма потребление кислорода тканями повышается, парциальное напряжение его в крови снижается, что приводит к компенсаторному усилению легочного дыхания. Таким образом, летом поглощение кислорода через легкие амфибий возрастает в несколько раз и намного превышает поглощение его через кожу. Поглощение же кислорода через кожу на протяжении всего года остается примерно постоянным в связи с постоянным содержанием кислорода в атмосферном воздухе, что обеспечивает постоянную скорость диффузии этого газа через поверхность кожи. Для обмена СО2 кожа более важна при всех температурах. Таким образом, при низких температурах кожа играет более важную роль, чем легкие, как в отдаче углекислого газа, так и в поглощении кислорода. При высоких температурах возрастает интенсивность легочного дыхания, обеспечивающая увеличение потребления организмом кислорода, тогда как кожное дыхание остается постоянным.
Кожа рептилий в отличие от кожи амфибий почти не проницаема для газов. Рептилии дышат легкими, и большинство их – наземные формы. Вместе с тем, настоящие морские змеи – действительно морские животные, и некоторые из них даже рождают в море живых детенышей. Они прекрасные пловцы и способны погружаться на глубину до 20 м. При этом, когда морская змея Pelamis platurus находится под водой, она может поглощать кислород через кожу с интенсивностью, доходящей до 33% от стандартного уровня, и выделять до 9% от обычного количества СО2. Несмотря на то, что у этой змеи главным органом газообмена служат легкие, кожа также оказывает существенную помощь, когда змея преследует мальков рыб.
Рис. Фото морской змеи Pelamis platurus, которая способна при нахождении под водой поглощать кислород через кожу.
Роль кожи в дыхании у млекопитающих ничтожно мала: поглощение кислорода через кожу едва измеримо, а выделение СО2 составляет менее 1% от количества, выводимого через легкие. В то же время у летучих мышей относительная поверхность кожи намного больше, чем у других млекопитающих; широкие, тонкие, лишенные волосяного покрова перепонки крыльев у них сильно васкуляризованы и могли бы участвовать в газообмене. Через эти перепонки выделяется некоторое количество углекислого газа. Так, при 18°С у летучей мыши Eptesicus fuscus через кожу крыльев выделяется 0,4% всей образующейся двуокиси кислорода. Эта доля возрастает с температурой воздуха, и при 27,5°С составляет 11,5% СО2. Однако поглощение кислорода через перепонки крыльев не настолько велико, чтобы оно имело какое-то значение, поскольку скорость диффузии кислорода между кровью и воздухом в 25 раз меньше, чем скорость диффузии углекислого газа.
Рис. Фото летучей мыши Eptesicus fuscus, у которой в выведении углекислого газа наружу может участвовать кожа
Дыхательные движения у наземных животных Для поступления в легкие наземных животных воздуха необходимо циклическое осуществление дыхательных движений. Существуют 2 основных способа, обеспечивающих поступление в легкие воздуха: ü нагнетательный насос (типичный для большинства амфибий) ü всасывающий насос (типичный для большинства рептилий, птиц и млекопитающих).
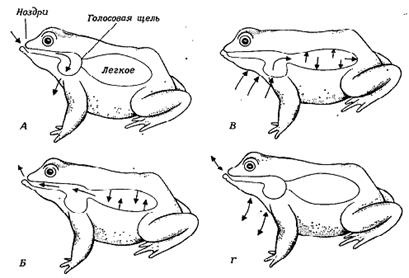
Сущность работы нагнетательного насоса состоит в следующем (см. рис.). Лягушка, чтобы наполнить легкие, вначале набирает воздух в ротовую полость (путем опускания ее дна). Затем она закрывает рот и ноздри и путем подъема дна ротовой полости обеспечивает повышение давления в ней, в результате чего воздух по градиенту давления переходит в легкие. Благодаря данному механизму наполнения легких лягушка может несколько раз подряд набирать новые порции воздуха, не позволяя ему выходить, в результате чего она способна раздуться до значительных размеров.
Рис. Дыхательный цикл у лягушки (Rana) (Gans, 1969). А – воздух набирается в ротовую полость путем опускания ее дна. В – ноздри закрываются, и воздух проталкивается в легкие. Г – до тех пор, пока голосовая щель остается сомкнутой, воздух удерживается в легких, и цикл может быть повторен, начиная с нового набирания воздуха в рот. Б – воздух выпускается из легких через верхнюю часть ротовой полости.
Рис. Фото лягушки и ящерицы Sauromalus после нескольких последовательных вдохов, не сопровождающихся выдохами
Нагнетание под действием положительного давления, аналогичное механизму амфибий, обнаружено и у некоторых рептилий. Так, ящерица Sauromalus, обитающая в пустынях юго-западной части Северной Америки, часто прячется в трещинах скал, где, раздув свои легкие, она заклинивается так прочно, что ее невозможно вытащить. Единственный способ достать это животное, используемый индейцами, употребляющими его в пищу, – проколоть его заостренной палочкой. Поступление воздуха в легкие путем всасывания типично для большинства рептилий, а также птиц и млекопитающих. Для его осуществления необходима герметично замкнутая грудная полость, периодическое увеличение объема которой, приводит к понижению давления в грудной полости и легких (до уровня, меньшего атмосферного) и, как следствие, поступлению в легкие воздуха из окружающей среды по воздухоносным путям по градиенту давления. При этом у млекопитающих увеличение объема грудной полости достигается в результате сокращения диафрагмы и межреберных мышц, приводящему к увеличению объема грудной полости и понижению давления в ней, что обуславливает понижение давление и в самих легких, и как следствие втягивание в них воздуха. Диафрагма млекопитающих представляет собой грудно-брюшную перегородку, в центре которой расположен сухожильный центр. От сухожильного центра радиально отходят мышечные волокна, прикрепляющиеся к грудине, ребрам и позвоночнику. Птицы имеют перепончатую диафрагму, прикрепленную к стенкам тела с помощью мышц, но по своей функции она отличается от диафрагмы млекопитающих.
Особенности строения легких у позвоночных в связи с разным уровнем их организации По мере перехода от одного класса позвоночных к другому по восходящей линии имеет место усложнение структурной организации легких. Так, у амфибий легкое представляет собой одиночный мешок, подразделенный несколькими гребнями, увеличивающими площадь поверхности. Легкие млекопитающих разделены на множество гораздо более мелких мешочков – альвеол (до 500 мкм в диаметре), которые во много раз увеличивают поверхность, пригодную для газообмена. Так, 1 см3 легочной ткани лягушки имеет общую газообменную поверхность 20 см2, тогда как у человека – 300 см2. При этом у млекопитающих легкие занимают около 6% объема тела независимо от его веса. Это означает, что мелкие млекопитающие, у которых интенсивность обмена высока, получают достаточно кислорода через легкие той же относительной величины, что и у крупных животных. У ныряющих животных, таких как дельфин, ламантин и кит, относительный объем легких такой же, как и у мелких млекопитающих, поскольку время пребывания их под водой не определяется запасами кислорода в легких. Гораздо более высокая площадь газообменной поверхности легких у гомойтермных животных обусловлена более высоким уровнем метаболизма у них, требующим большей интенсивности поглощения кислорода. Интенсивность дыхания у млекопитающих точно подстраивается под потребности организма в кислороде, определяемого уровнем его метаболизма, зависящего от функциональной активности. Так, у человека в покое дыхательный объем составляет около 500 мл, из которых 150 мл заполняет воздухоносные пути (анатомически мертвое пространство), а 350 мл обеспечивает вентиляцию альвеол (иными словами, в покое объем анатомически мертвого пространства составляет около 1/3 от дыхательного объема). При этом после спокойного выдоха в легких остается около 2400 мл воздуха (т.н. функциональная остаточная емкость легких, включающая остаточный объем и резервный объем выдоха). Таким образом, при спокойном дыхании в момент каждого вдоха в альвеолах легких обновляется 1/7 часть воздуха. При интенсификации дыхания в момент, например, физической активности, человек вдыхает за один раз около 3000 мл воздуха, из которых 2850 мл идет на вентиляцию альвеол, а 150 мл – на вентиляцию воздухоносных путей. Иными словами, при повышении функциональной активности объем воздуха, идущий на вентиляцию анатомически мертвого пространства, составляет 1/20 дыхательного объема. Таким образом, в покое анатомически мертвое пространство составляет значительную часть дыхательного объема, а при нагрузке — относительно несущественную. Однако при усилении вентиляции альвеол затраты на дыхание несколько возрастают. Так, если вентиляция легких у человека увеличивается до 10 л/мин, эти затраты достигают 1 мл кислорода на 1 л воздуха, а при очень интенсивной вентиляции (50 л/мин) они возрастают до 2 мл на 1 л воздуха. По другим оценкам, максимальные затраты на дыхание при нагрузке не превышают 3% от всего поглощенного кислорода, т.е. несколько меньше приведенных значений. Затраты на дыхание у других млекопитающих находятся в том же диапазоне, что и у человека. Для сравнения затраты на дыхание у рыб, которым приходится перемещать гораздо более тяжелую и вязкую среду (воду), тоже невысоки: находятся примерно в том же диапазоне, что и у млекопитающих (несколько процентов от общего потребления кислорода), что обусловлено однонаправленным током воды в жабрах. Без однонаправленного тока воды затраты на вентиляцию, вероятно, не могли бы быть столь малыми (могли бы достигать 30-50%). Пузыреобразная форма и большая кривизна альвеол в легких позвоночных обуславливает высокое поверхностное натяжение во влажных внутренних их поверхностях, которое стремится заставить альвеолы спасться. Но при этом тенденция к сжатию альвеол сводится к минимуму благодаря присутствию на стенках альвеол веществ, сильно уменьшающих поверхностное натяжение. Эти вещества — фосфолипиды, и за свое влияние на поверхностное натяжение они получили название сурфактантов. Сурфактанты обнаружены в легких всех позвоночных — млекопитающих, птиц, рептилий и амфибий. Количество сурфактантов в легких позвоночных, по-видимому, всегда превышает тот минимум, который требуется для покрытия всей легочной поверхности мономолекулярным слоем (см. рис.).
Рис. Зависимость между площадью поверхности легких и количеством сурфактанта, которое можно экстрагировать из легких различных позвоночных. Во всех случаях это количество больше теоретического минимума, необходимого для покрытия легочной поверхности мономолекулярным слоем. (Clements et al., 1970.)
Особенности дыхания некоторых рыб, способных дышать не только в воде, но и в воздухе
Несмотря на то, что большинство морских и пресноводных рыб способны только к водному дыханию и при изъятии их из воды погибают от удушья (в связи с тем, что жабры спадаются и не могут эффективно извлекать кислород из воздуха), существует ряд рыб, способных переходить на воздушное дыхание в случае, когда концентрация кислорода в воде низка. Относительно немногие рыбы зависят от воздуха в такой степени, что погибают, если их удерживать под водой. Существует две главные экологические причины для использования у рыб воздушного дыхания в качестве дополнительного или единственного: 1) нехватка кислорода в воде, 2) периодические засухи.
Двоякодышащая рыба, например, в засушливые периоды зарывается глубоко в ил, окружает себя капсулой и остается неактивной до следующего влажного периода (т.е. временно впадает в состояние анабиоза). Большая часть воздуходышащих рыб — обитатели тропических пресных водоемов и эстуариев. Истинно морские двоякодышащие виды если и встречаются, то их очень мало. При этом пресные воды с дефицитом О2 гораздо чаще встречается в тропиках, чем в умеренных зонах. Это объясняется тем, что в тропических водах много разлагающегося органического материала, температура высока и ускоряет деятельность бактерий; небольшие водоемы часто сильно затенены нависающими джунглями (что снижает фотосинтез и образование кислорода в воде); температура мало изменяется на протяжении суток, в связи с чем тепловая конвекция, которая могла бы доставить богатую кислородом поверхностную воду к глубинным слоям, очень слаба. В связи с отмеченными особенностями пресных водоемов тропических зон в процессе эволюции у рыб выработалась особенность использовать для дыхания воздух. Вместе с тем, не все дышащие воздухом рыбы обитают в тропиках. Хорошо известная ильная рыба (Amia calva) встречается в северной части США даже в тех местах, где озера зимой замерзают. В такие периоды эта рыба легко обходится без воздуха, так как низкая температура снижает интенсивность обменных процессов и соответственно потребление кислорода. Несмотря на то, что жабры рыб мало приспособлены для дыхания в воздухе: у них нет необходимой жесткости, и они имеют тенденцию слипаться, они все же могут извлекать из воздуха некоторое количество кислорода. Наряду с жабрами, любая другая влажная поверхность тоже будет вносить некоторый вклад в газообмен, если эта поверхность контактирует с воздухом и снабжается кровью. Так, некоторый газообмен всегда может осуществляться через кожу и поверхность ротовой полости. Кроме того, у некоторых рыб существуют другие, анатомически более специализированные органы, помогающие газообмену. Органы, которые обычно используются для дыхания воздухом, — это жабры, кожа, ротовая и оперкулярные полости, желудок, кишечник, плавательный пузырь и легкие. Большинство двоякодышащих рыб – это высшие костные рыбы (лучеперые — Actinopterygii), способные дышать воздухом при недостатке кислорода в воде или пересыхании водоемов, используя при этом жабры или какие-то другие альтернативные органы (ротовую полость, желудок, кишечник, плавательный пузырь и другие), но в то же время основным типом дыхания для них является водное дыхание с участием жабр. Таким образом, основным для этих рыб является водное дыхание, но в то же время при неблагоприятных условиях они могут переходить к воздушному дыханию. Среди таких рыб можно назвать следующие: ü ильная рыба (Amia) – представитель примитивной группы Ноlostei, использует для дыхания плавательный пузырь, обитает в пресных водах Северной Америки, вобластях, где озера всю зиму покрыты льдом; принадлежит к примитивной группе костных рыб
ü многопер (Polypterus), имеет легкие, но не является настоящей двоякодышащей рыбой, обитает в пресных водах Африки, принадлежит к примитивной группе костных рыб
Поиск по сайту: |
































 краб Cardiosoma
краб Cardiosoma