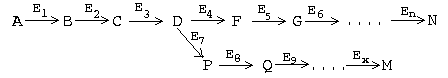|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
АВТОНОМНАЯ САМОРЕГУЛЯЦИЯ ФЕРМЕНТАТИВНЫХ ПРОЦЕССОВ ⇐ ПредыдущаяСтр 3 из 3
Существуют различные механизмы регуляции процессов, протекающих в организме. 3 уровня регуляции: 1) Самый молодой уровень филогенетически - НЕЙРОГУМОРАЛЬНАЯ регуляция (с участием центральной нервной системы, классических гормонов и гормонов местного действия) 2) РЕГУЛЯЦИЯ НА ГЕНЕТИЧЕСКОМ УРОВНЕ - изменение скорости биосинтеза белка. 3) Филогенетически наиболее старый уровень - АВТОНОМНАЯ САМОРЕГУЛЯЦИЯ ФЕРМЕНТОВ. АВТОНОМНАЯ САМОРЕГУЛЯЦИЯ - это регуляция, которая происходит благодаря только самим участникам реакции, то есть за счет фермента, его субстрата (или субстратов) и/или продуктов деятельности данного фермента. Фермент не только работает, но еще и сам себя регулирует. Механизмы автономной саморегуляции очень многочисленны, но построены на двух основных принципах. 1) Механизмы, основанные на кинетических свойствах фермента, количественно характеризуемых Км и Vmax - это механизмы КИНЕТИЧЕСКОГО ТИПА. 2) Второй принцип связан с аллостерическими свойствами фермента, то есть со способностью фермента угнетаться или активироваться под действием субстрата и/или продуктов. Это механизмы АЛЛОСТЕРИЧЕСКОГО ТИПА. Такие аллостерические механизмы есть не у каждого фермента. Они обычно накладываются на механизмы кинетического типа и определяют особенности регуляции данного фермента. E А --------> B
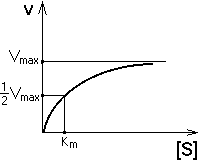
Кинетика данной ферментативной реакции описывается уравнением Михаэлиса-Ментен и кривой, изображенной на рисунке.
Если в клетку поступает мало субстрата, то он медленно и расщепляется. При изменении скорости поступления субстрата в клетку фермент работает так, чтобы поддержать концентрацию субстрата в клетке около Км. На этот механизм могут накладываться механизмы аллостерического типа:
1. СУБСТРАТ или ПРОДУКТ - аллостерический ИНГИБИТОР своего фермента.
При чрезмерном поступлении субстрата в клетку скорость утилизации субстрата все больше будет замедляться. Так происходит, когда избыток продукта реакции опасен для клетки (опаснее, чем избыток субстрата).
2. СУБСТРАТ - аллостерический АКТИВАТОР своего фермента.
Кинетическая кривая имеет S-образный характер, то есть имеет 2 перегиба, как кривая диссоциации оксигемоглобина. В этом случае концентрация субстрата удерживается более эффективно на постоянном уровне и в более узком диапазоне, чем в предыдущем случае.
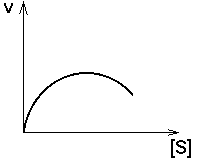
3. ПРОДУКТ реакции - аллостерический АКТИВАТОР своего фермента.
Кинетическая кривая имеет лавинообразный (взрывообразный) характер. С увеличением концентрации субстрата скорость реакции, как обычно, возрастает. Это приводит к накоплению продукта, который активирует свой фермент. Это приводит к накоплению продукта, который активирует фермент, в результате продукт накапливается еще быстрее, а фермент активируется еще сильнее. Скорость реакции становится очень большой, и реакция протекает мгновенно до полного расщепления субстрата. Примеры: реакция образования фибрина из фибриногена; система иммунохимической защиты организма.
4. ОДИН СУБСТРАТ - ДВА ФЕРМЕНТА И ДВА ПРОДУКТА.
В общем случае кинетические кривые этих двух реакций не совпадают.
При малых концентрациях субстрата "А" будет преимущественно протекать 1-я реакция, то есть наибольшая часть вещества "А" будет превращаться в "В", а меньшая часть - превращаться в "С". При увеличении концентрации "А" все больше субстрата станет превращаться в "С" по второму пути. При некоторой концентрации "А" (в точке пересечения кривых) скорости обеих реакций сравняются. В этой точке половина вещества "А" будет превращаться в "В", а вторая половина - в "С". Если концентрацию "А" повышать дальше, то начнет преобладать второй путь превращений - в вещество "С". Такой механизм регуляции встречается, когда опасно накопление в клетке не только избытка вещества "В", но и избытка субстрата "А". И поэтому клетка при высоких концентрациях "А" направляет его на образование безопасного продукта С. Путь из "А" в "С" - это резервный путь метаболизма (альтернативный путь). Если на такой механизм накладываются аллостерические механизмы регуляции, то они могут стать более точными. Например, если продукт "В" - аллостерический ингибитор своего фермента Е1 или аллостерический активатор фермента Е2, то переключение на резервный (альтернативный) путь будет происходить более четко.
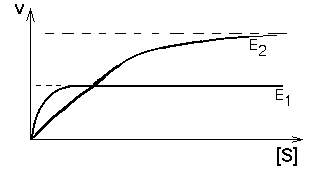
5. ОДИН СУБСТРАТ, ДВА ФЕРМЕНТА И ОДИН ПРОДУКТ Одна и та же химическая реакция в организме может катализироваться разными белками-ферментами. В таком случае они называются изоферментами - это разные молекулярные формы одного и того же фермента. Они могут отличаться обычно очень незначительно. Например, в молекуле один или несколько аминокислотных остатков могут быть заменены другими. Но этого достаточно, чтобы возникли различия в значениях ИЭТ, оптимальном значении рН для действия фермента (рН-оптимум), и в субстратной специфичности, и в величинах Vmax и Km. Называют такие ферменты, как правило, одинаково, но добавляют к названию номер или иное дополнение (для идентификации). И в этом случае, если определять кинетику ферментативных реакций изоферментов, то кривые будут отличаться друг от друга. Если ферменты находятся в клетках разного типа, то изоферменты будут определять специфику метаболизма своих клеток.
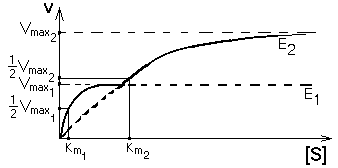
Но если изоферменты находятся в одной клетке, то их кинетические кривые будут сливаться в одну общую кривую, которая имеет двухступенчатую форму. Такая система регуляции работает при разных концентрациях субстрата, которые изменяются в очень широких пределах. Наличие двух изоферментов позволяет успешно превращать субстрат и при малых, и при больших концентрациях субстрата. Характер суммарной кинетической кривой приведен на рисунке.
Конкретным примером такого типа регуляции служит пара ферментов - гексокиназа и глюкокиназа в печени. Км гексокиназы составляет около 0,02 ммоль/л. В клетках печени кроме гексокиназы есть глюкокиназа, имеющая Км=20 ммоль/л. В норме концентрация глюкозы в крови колеблется в пределах от 3,9 до 6,1 ммоль/л. Так как Км гексокиназы в 100 раз меньше, чем реальная концентрация глюкозы в клетке, то гексокиназа всегда работает со скоростью, близкой к Vmax. Благодаря высокому сродству к своему субстрату - глюкозе именно гексокиназа обеспечивает связывание глюкозы в печени в период голодания. На высоте пищеварения, когда концентрация глюкозы в системе воротной вены может составлять десятки ммоль/л, гексокиназа уже не справляется с превращением такого большого количества глюкозы. Наибольшая часть ее превращается глюкокиназой - мощным ферментом, Км которого достигает 20 ммоль/л. Поэтому глюкоза эффективно связывается клетками печени и в период голодания (за счет работы гексокиназы), и на высоте пищеварения (с помощью глюкокиназы). Благодаря такому механизму уровень глюкозы в крови, оттекающей от печени, поддерживается на нужном уровне при любых колебаниях ее концентрации в системе воротной вены. НЕРАЗВЕТВЛЕННЫЕ МУЛЬТИФЕРМЕНТНЫЕ СИСТЕМЫ В большинстве случаев тот или иной биохимический процесс катализируется не одним ферментом, а целой совокупностью ферментов, каждый из которых катализирует свой этап в длинной последовательности отдельных реакций этого процесса. Совокупность ферментов, катализирующих последовательные реакции единого процесса, называется мультиферментной системой. E1 E2 E3 E4 En А---->В----->C----->D------>....L----->M
1. В такой системе концентрация любого промежуточного метаболита от "В" и до "L" будет поддерживаться постоянной даже в условиях значительных колебаний скорости всего процесса в целом. Концентрация любого метаболита зависит только от соотношения кинетических констант 2-х ферментов - того, который ведет к образованию этого метаболита, и того, который подвергает его следующему превращению. Например, концентрация вещества "С" зависит от кинетических констант ферментов Е» и Е3. Оба этих фермента, как и вся цепь, могут ускорять свою работу, но концентрация "С" будет оставаться неизменной. Эти механизмы регуляции мультиферментных систем поддерживают постоянство состава внутренней среды организма. 2. В большинстве случаев величины Vmax у всех ферментов данной системы различны. Значит, у какого-то из ферментов Vmax меньше Vmax любого другого фермента. Такой фермент (с низкой Vmax) называется ЛИМИТИРУЮЩИМ ферментом данной мультиферментной системы. Именно этот фермент определяет скорость процесса в целом. Как правило, лимитирующими являются те ферменты, которые находятся в начале данной мультиферментной системы. Как правило, эти ферменты подвергаются воздействию регуляторных воздействий со стороны аллостерических эффекторов (в рамках автономной саморегуляции). В таких мультиферментных системах аллостерическими эффекторами бывают часто не только продукт или субстрат данной реакции, но и другие метаболиты - в особенности конечные продукты (М). Если конечный продукт оказывает активирующее действие, то этот механизм называется положительной обратной связью и процесс протекает взрывообразно - до полного исчерпания субстрата в системе. Если конечный продукт - ингибитор лимитирующего фермента - то говорят об отрицательной обратной связи, и при увеличении концентрации конечного продукта скорость процесса будет замедляться. Если лимитирующий фермент является одновременно и регуляторным, то его называют КЛЮЧЕВЫМ ферментом данной мультиферментной цепи.
РАЗВЕТВЛЕННЫЕ МУЛЬТИФЕРМЕНТНЫЕ СИСТЕМЫ. Часто мультиферментные цепи являются разветвленными. В таких системах ключевые ферменты обычно предшествуют участкам ветвления, а сразу за разветвлением находятся ПУНКТЫ ВТОРИЧНОГО КОНТРОЛЯ, то есть такие ключевые ферменты, которые регулируют скорость реакций в своей ветви:
В этом примере Е1 - ключевой фермент, а Е4 и Е7 - пункты вторичного контроля.
[j1]
Поиск по сайту: |