|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Локализация функций в коре большого мозга
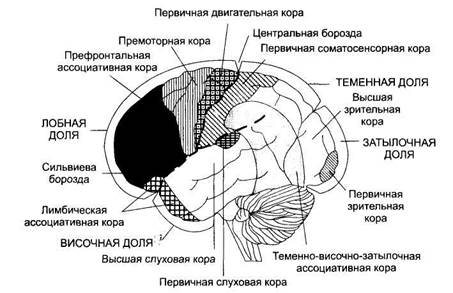
Общая характеристика.В определенных участках коры большого мозга сосредоточены преимущественно нейроны, воспринимающие один вид раздражителя: затылочная область — свет, височная доля — звук и т.д. Однако после удаления классических проекционных зон (слуховых, зрительных) условные рефлексы на соответствующие раздражители частично сохраняются. Согласно теории И. П. Павлова в коре большого мозга имеются «ядро» анализатора (корковый конец) и «рассеянные» нейроны по всей коре. Современная концепция локализации функций базируется на принципе многофункциональности (но не равноценности) корковых полей. Свойство мультифункциональности позволяет той или иной корковой структуре включаться в обеспечение различных форм деятельности, реализуя при этом основную, генетически присущую ей функцию (О. С. Адрианов). Степень мультифункциональности различных корковых структур неодинакова. В полях ассоциативной коры она выше. В основе мультифункциональности лежит многоканальность поступления в кору мозга афферентного возбуждения, перекрытия афферентных возбуждений, особенно на таламическом и корковом уровнях, модулирующее влияние различных структур, например неспецифических ядер таламуса, базальных ганглиев на корковые функции, взаимодействие корково-подкорковых и межкорковых путей проведения возбуждения. С помощью микроэлектродной техники удалось зарегистрировать в различных областях коры большого мозга активность специфических нейронов, отвечающих на стимулы только одного вида раздражителя (только на свет, только на звук и т.п.), т.е. имеется множественное представительство функций в коре большого мозга. В настоящее время принято подразделение коры на сенсорные, двигательные и ассоциативные (неспецифические) зоны (области). Сенсорные зоны коры.Сенсорная информация поступает в проекционную кору, корковые отделы анализаторов (И.П.Павлов). Эти зоны расположены преимущественно в теменной, височной и затылочной долях. Восходящие пути в сенсорную кору поступают в основном от релейных сенсорных ядер таламуса. Первичные сенсорные зоны — это зоны сенсорной коры, раздражение или разрушение которых вызывает четкие и постоянные изменения чувствительности организма (ядра анализаторов, по И. П. Павлову). Они состоят преимущественно из моносенсорных нейронов и формируют ощущения одного качества. В первичных сенсорных зонах обычно имеется четкое пространственное (топографическое) представительство частей тела, их рецепторных полей. Первичные проекционные зоны коры состоят главным образом из нейронов IV афферентного слоя, для которых характерна четкая топическая организация. Значительная часть этих нейронов обладает высочайшей специфичностью. Так, например, нейроны зрительных областей избирательно реагируют на определенные признаки зрительных раздражителей: одни — на оттенки цвета, другие — на направление движения, третьи — на характер линий (край, полоса, наклон линии) и т.п. Однако следует отметить, что в первичные зоны отдельных областей коры включены также нейроны мультисенсорного типа, реагирующие на несколько видов раздражителей. Кроме того, там же имеются нейроны, реакция которых отражает воздействие неспецифических (лимбико-ретикулярных, или модулирующих) систем. Вторичные сенсорные зоны расположены вокруг первичных сенсорных зон, менее локализованы, их нейроны отвечают преимущественно на действие двух раздражителей, т.е. они бисенсорны. Третичная сенсорная зона — ассоциативная кора (см. ниже). Локализация сенсорных зон.Важнейшей сенсорной областью является теменная доля (постцентральной извилины и соответствующая ей часть парацентральной дольки на медиальной поверхности полушарий). Эту зону обозначают как соматосенсорную область I. Здесь имеется проекция кожной чувствительности противоположной стороны тела от тактильных, болевых, температурных рецепторов, интероцептивной чувствительности и чувствительности опорно-двигательного аппарата — от мышечных, суставных, сухожильных рецепторов (рис. 2). Кроме соматосенсорной области I выделяют соматосенсорную область II меньших размеров, расположенную на границе пересечения центральной борозды с верхним краем височной доли, в глубине латеральной борозды. Точность локализации частей тела здесь выражена в меньшей степени. Хорошо изученной первичной проекционной зоной является слуховая кора (поля 41, 42), которая расположена в глубине латеральной борозды (кора поперечных височных извилин Гешля). К проекционной коре височной доли относится также центр вестибулярного анализатора в верхней и средней височных извилинах. В затылочной доле расположена первичная зрительная область (кора части клиновидной извилины и язычковой дольки, поле 17). Здесь имеется топическое представительство рецепторов сетчатки. Каждой точке сетчатки соответствует свой участок зрительной коры, при этом зона желтого пятна имеет сравнительно большую зону представительства. В связи с неполным перекрестом зрительных путей в зрительную область каждого полушария проецируются одноименные половины сетчатки. Наличие в каждом полушарии проекции сетчатки обоих глаз является основой бинокулярного зрения. Около поля 17 расположена кора вторичной зрительной области (поля 18 и 19). Нейроны этих зон полисенсорны и отвечают не только на световые, но и на тактильные и слуховые раздражители. В данной зрительной области происходит синтез различных видов чувствительности, возникают более сложные зрительные образы и их опознание. Из первичных и вторичных сенсорных зон информация передается для дальнейшей ее обработки в ассоциативную кору (третичная сенсорная зона), после чего инициируется (в случае необходимости) поведенческая реакция с обязательным участием двигательной коры.
Рис. 2. Схема чувствительного и двигательного гомункулусов (по У. Пенфилду, Т. Расмуссену). Разрез полушарий во фронтальной плоскости: а — проекция общей чувствительности в коре постцентральной извилины; б— проекция двигательной системы в коре прецентральной извилины
Двигательные зоны коры.Выделяют первичную и вторичную моторные зоны. В первичной моторной зоне (прецентральная извилина лобных долей, поле 4) расположены нейроны, иннервирующие мотонейроны мышц лица, туловища и конечностей. В ней имеется четкая топографическая проекция мышц тела (см. рис. 2). Основной закономерностью топографического представительства является то, что регуляция деятельности мышц, обеспечивающих наиболее точные и разнообразные движения (речь, письмо, мимика), требует участия больших по площади участков двигательной коры. Раздражение первичной моторной коры вызывает сокращение мышц противоположной стороны тела (для мышц головы сокращение может быть билатеральным). При поражении этой корковой зоны утрачивается способность к тонким координированным движениям конечностями, особенно пальцами рук. Вторичная моторная зона (поле 6) расположена как на латеральной поверхности полушарий, впереди прецентральной извилины (премоторная кора), так и на медиальной поверхности, соответствующей коре верхней лобной извилины (дополнительная моторная область). Вторичная двигательная кора в функциональном плане имеет главенствующее значение по отношению к первичной двигательной коре, осуществляя высшие двигательные функции, связанные с планированием и координацией произвольных движений. Здесь в наибольшей степени регистрируется медленно нарастающий отрицательный потенциал готовности, возникающий примерно за 1 с до начала движения. Кора поля 6 получает основную часть импульсации от базальных ядер и мозжечка, участвует в перекодировании информации о плане сложных движений. Раздражение коры поля 6 вызывает сложные координированные движения, например поворот головы, глаз и туловища в противоположную сторону, содружественные сокращения сгибателей или разгибателей на противоположной стороне. В премоторной коре расположены двигательные центры, связанные с социальными функциями человека: центр письменной речи в заднем отделе средней лобной извилины (поле 6), центр моторной речи Брока в заднем отделе нижней лобной извилины (поле 44), обеспечивающие речевой праксис, а также музыкальный моторный центр (поле 45), обеспечивающий тональность речи, способность петь. Нейроны двигательной коры получают афферентные входы через таламус от мышечных, суставных и кожных рецепторов, от базальных ядер и мозжечка. Основными эфферентными выходами двигательной коры являются пути на стволовые и спинальные моторные центры. Основные доли коры большого мозга представлены на рис. 3. Ассоциативные области (зоны) коры(неспецифическая, межсенсорная, межанализаторная кора). Они включают участки новой коры большого мозга, которые расположены вокруг проекционных зон и рядом с двигательными зонами, но не выполняют непосредственно чувствительных или двигательных функций, поэтому им нельзя приписывать преимущественно сенсорные или двигательные функции. Нейроны этих зон обладают большими способностями к обучению. Границы этих областей обозначены недостаточно четко. Ассоциативная область коры является филогенетически наиболее молодой частью новой коры, получившей наибольшее развитие у приматов и человека. У человека она составляет около 50 % всей коры или 70 % неокортекса. Термин «ассоциативная кора» возник в связи с существовавшим представлением о том, что эти зоны за счет проходящих через них кортико-кортикальных соединений связывают двигательные зоны и одновременно служат субстратом высших психических функций. Основными ассоциативными зонами коры являются: теменно-височно-затылочная, префронтальная кора лобных долей и лимбическая ассоциативная зона.
Рис. 3. Четыре основные доли коры головного мозга: лобная, височная, теменная и затылочная (вид сбоку). В них расположены первичная двигательная и сенсорная области, двигательные и сенсорные области более высокого порядка (второго, третьего и т.д.), ассоциативная (неспецифичная) кора
Нейроны ассоциативной коры (ее называют также третичной сенсорной зоной) являются полисенсорными: они отвечают, как правило, не на один (как нейроны первичных сенсорных зон), а на несколько раздражителей, т.е. один и тот же нейрон может возбуждаться при раздражении слуховых, зрительных, кожных и других рецепторов. Полисенсорность нейронов ассоциативной коры создается кортико-кортикальными связями с разными проекционными зонами, связями с ассоциативными ядрами таламуса. В результате этого ассоциативная кора представляет собой своеобразный коллектор различных сенсорных возбуждений и участвует в интеграции сенсорной информации и в обеспечении взаимодействия сенсорных и моторных областей коры. Работа ассоциативных областей коры мозга необходима не только для успешного синтеза и дифференцирования (избирательного различения) воспринимаемых человеком раздражителей, но и для перехода к уровню их символизации, т.е. для оперирования значениями слов и использования их для отвлеченного мышления, для синтетического характера восприятия. С 1949 г. широкую известность получила гипотеза Д. Хебба, постулирующая в качестве условия синаптической модификации совпадение пресинаптической активности с разрядом постсинаптического нейрона, поскольку не всякая активность синапса ведет к возбуждению постсинаптического нейрона. На основании данной гипотезы можно предположить, что отдельные нейроны ассоциативных зон коры связаны разнообразными путями и образуют клеточные ансамбли, выделяющие «подобразы», т.е. соответствующие унитарным формам восприятия. Эти связи, как отмечал Д. Хебб, настолько хорошо развиты, что достаточно активировать один нейрон, как возбуждается весь ансамбль. Аппаратом, выполняющим роль регулятора уровня бодрствования, а также осуществляющим избирательную модуляцию и актуализацию приоритета той или иной функции, является модулирующая система мозга, которую часто называют лимбико-ретикулярным комплексом, или восходящей активирующей системой. К нервным образованиям этого аппарата относятся лимбическая и неспецифические системы мозга с активирующими и инактивирующими структурами. Среди активирующих образований прежде всего выделяют ретикулярную формацию среднего мозга, задний гипоталамус, голубое пятно в нижних отделах ствола мозга. К инактивирующим структурам относят преоптическую область гипоталамуса, ядра шва в стволе мозга, фронтальную кору. Таламокортикальные системы.В настоящее время поталамокортикальным проекциям выделяют триосновные ассоциативные системы мозга: таламотеменную, таламолобную и таламовисочную. Таламотеменная система представлена ассоциативными зонами теменной коры, получающими основные афферентные входы от задней группы ассоциативных ядер таламуса. Теменная ассоциативная кора имеет эфферентные выходы на ядра таламуса и гипоталамуса, в моторную кору и ядра экстрапирамидной системы. Основными функциями таламотеменной системы являются гнозис и праксис. Под гнозисом понимают функцию различных видов узнавания: формы, величины, значения предметов, понимание речи, познание процессов, закономерностей и др. К гностическим функциям относится оценка пространственных отношений, например взаимного расположения предметов. В теменной коре выделяют центр стереогнозиса, обеспечивающий способность узнавания предметов наощупь. Вариантом гностической функции является формирование в сознании трехмерной модели тела («схемы тела»). Под праксисом понимают целенаправленное действие. Центр праксиса находится в надкраевой извилине левого полушария, он обеспечивает хранение и реализацию программы двигательных автоматизированных актов. Таламолобная система представлена ассоциативными зонами лобной коры, имеющими основной афферентный вход от ассоциативного медиодорзального ядра таламуса, других подкорковых ядер. Основная роль лобной ассоциативной коры сводится к инициации базовых системных механизмов формирования функциональных систем целенаправленных поведенческих актов (П. К. Анохин). Префронтальная область играет главную роль в выработке стратегии поведения. Нарушение этой функции особенно заметно, когда необходимо быстро изменить действие и когда между постановкой задачи и началом ее решения проходит некоторое время, т.е. успевают накопиться раздражители, требующие правильного включения в целостную поведенческую реакцию. Таламовисочная система представляет некоторые ассоциативные центры, например стереогнозиса, праксиса, включающие в себя и участки височной коры. В височной коре расположен слуховой центр речи Вернике, находящийся в задних отделах верхней височной извилины левого полушария. Этот центр обеспечивает речевой гнозис: распознание и хранение устной речи как собственной, так и чужой. В средней части верхней височной извилины находится центр распознания музыкальных звуков и их сочетаний. На границе височной, теменной и затылочной долей находится центр чтения, обеспечивающий распознание и хранение образов. Существенную роль в формировании поведенческих актов играет биологическое качество безусловной реакции, а именно ее значение для сохранения жизни. В процессе эволюции это значение было закреплено в двух противоположных эмоциональных состояниях — положительном и отрицательном, которые у человека составляют основу его субъективных переживаний: удовольствия и неудовольствия, радости и печали. Во всех случаях целенаправленное поведение строится в соответствии с эмоциональным состоянием, возникшим при действии раздражителя. Во время поведенческих реакций отрицательного характера напряжение вегетативных компонентов, особенно сердечнососудистой системы, в отдельных случаях, чаще в непрерывных так называемых конфликтных ситуациях, может достигать большой силы, что вызывает нарушение их регуляторных механизмов (вегетативные неврозы). В этой части книги рассмотрены основные вопросы аналитико-синтетической деятельности мозга, которые позволят перейти в последующих главах к изложению частных вопросов физиологии сенсорных систем и высшей нервной деятельности.
Поиск по сайту: |