|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Структурно-функциональная организация новой коры большого мозга
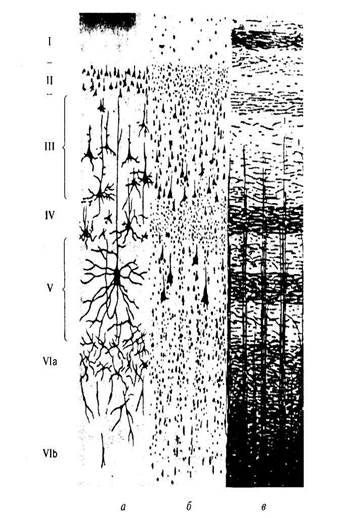
Кора большого мозга представляет собой многослойную нейронную ткань с множеством складок общей площадью в обоих полушариях примерно 2200 см2, что соответствует квадрату со сторонами 47x47 см, ее объем равен 40% массы головного мозга, толщина колеблется от 1,3 до 4,5 мм, а общий объем составляет 600 см3. В состав коры большого мозга входит 109—1010 нейронов и множество глиальных клеток, общее число которых пока неизвестно. Новая кора (неокортекс) — это слой серого вещества общей площадью 1100—1900 см2, покрывающий большие полушария и составляющий около 72 % всей площади коры. Слои новой коры большого мозга. Внаправлении с поверхности вглубь коры различают шесть (1 — VI) горизонтальных слоев (рис. 1). I. Молекулярный слой, имеющий мало клеток, но содержащий большое количество ветвящихся восходящих дендритов пирамидных клеток, на которых образуют синапсы волокна, приходящие от ассоциативных и неспецифических ядер таламуса и регулирующие уровень возбудимости коры. II. Наружный зернистый слой содержит в основном звездчатые и частично малые пирамидальные клетки. Волокна его клеток расположены преимущественно вдоль поверхности коры, образуя кортико-кортикальные связи. III. Наружный пирамидный слой сформирован в основном из пирамидальных клеток средней величины, а их аксоны образуют кортико-кортикальные ассоциативные связи, как и зернистые клетки IIслоя. IV. Внутренний зернистый слой образован звездчатыми клетками, на которых образуют синапсы волокна, идущие от нейронов специфических ядер таламуса и метаталамуса и несущие информацию от рецепторов сенсорных систем. V. Внутренний пирамидный слой представлен средними и крупными пирамидными клетками, при этом гигантские пирамидные клетки Беца расположены в двигательной коре. Их аксоны образуют эфферентные кортикобульбарный и кортикоспинальный двигательные пути. VI. Слой полиморфных клеток, аксоны которых образуют корти-коталамические пути. В I и IV слоях новой коры происходят восприятие и обработка поступающих сигналов. Нейроны II и IIIслоев осуществляют кортикокортикальные ассоциативные связи. Нейроны V и VI слоев формируют нисходящие пути.
Рис. 1. Полусхематичное изображение слоев коры головного мозга: а — основные типы нервных клеток (окраска по Гольджи); б — тела нейронов (окраска по Нисслю); в — общее расположение волокон (миелиновых оболочек). Слои пронумерованы от поверхности вглубь. Приведены две наиболее распространенные системы нумерации (объяснение в тексте)
Нейронные колонки.В коре мозга имеются функциональные объединения нейронов, расположенные в цилиндрике диаметром 0,5—1,0 мм, включающем все слои коры и содержащем несколько сотен нейронов (нейронные колонки). Об этом в частности свидетельствуют электрофизиологические исследования (В. Маунткасл, 1957) с погружением микроэлектродов перпендикулярно поверхности соматосенсорной коры. При этом все встречаемые на пути нейроны отвечают на раздражитель только одного вида (например, тактильный). При погружении электрода под углом на его пути попадаются нейроны разной сенсорности. Колонки обнаружены в моторной коре и различных зонах сенсорной коры. Нейроны коры могут осуществлять саморегуляцию по типу возвратного торможения. Соседние нейронные колонки могут частично перекрываться и взаимодействовать друг с другом по механизму латерального торможения. Нейронные модули.Несколько вертикальных колонок объединяются в единую крупную модульную колонку, получая и перерабатывая большой объем информации. Многократное поступление информации в корковые модули и ее циркуляция в замкнутых цепях обеспечивают электрические и химические изменения в нейронах и синапсах, необходимые для организации долговременной памяти. Отдельный нейрон получает информацию от множества других нейронов и посылает к ним импульсы. В результате происходит объединение индивидуальных нейронов в системы, обладающие новыми свойствами, отсутствующими у входящих в их состав единиц. Характерной чертой таких систем является то, что активность каждого составного элемента в них определяется не только влияниями, поступающими по прямым афферентным путям каждого элемента, но и состоянием других элементов системы. Системность в работе нервных образований возникает тогда, когда деятельность каждой нервной клетки оказывается функцией не только непосредственно поступившего к ней сигнала, но и функцией тех процессов, которые происходят в остальных клетках нервного центра (П. Г. Костюк). Обширные связи различных областей мозга между собой делятся условно на закрепленные (запрограммированные генетически) и развивающиеся в результате сенсорного притока и моторного опыта. Вследствие этого в мозге помимо жестко закрепленных организуются распределенные системы получения и обработки информации, системы, участвующие в обучении. Любой модуль может входить в любую систему обработки информации. Распределенные системы мозга анализируют информацию, поступающую из внешнего мира и внутренней среды организма. Хранится информация с помощью механизмов памяти. Распределенные системы имеют важное значение в формировании потребностей, мотиваций и эмоций. По своим биофизическим свойствамнейроны коры сходны с другими нервными клетками. Потенциал покоя пирамидных клеток составляет от -50 до -80 мВ, амплитуда их потенциала действия — 60 —100 мВ при длительности 0,5 — 2 мс. Потенциалы действия в нейронах коры возникают в области аксонного холмика и распространяются не только по аксону, но также по телу и дендритам клетки (по крайней мере, по проксимальным). Поскольку вслед за потенциалом действия в этих нейронах не возникает заметной следовой гиперполяризации, частота их импульсации достигает 100 Гц. В нейронах коры даже у бодрствующих животных частота импульсации, вызванной постсинаптическими потенциалами, низкая, обычно меньше 10 Гц, а нередко не достигает и 1 Гц. Потенциалы покоя клеток коры обычно колеблются в диапазоне на 3 — 10 мВ ниже порогового уровня (Екр), т.е. пороговый потенциал (∆v) колеблется и составляет 3 — 10 мВ. Кортикализация функций— это возрастание в филогенезе роли коры большого мозга в регуляции функций организма, в подчинении себе нижележащих отделов ЦНС, в обеспечении психической деятельности организма. Например, регуляция локомоторных двигательных функций (прыжки, ходьба, бег) и выпрямительных рефлексов у низших позвоночных полностью обеспечивается стволом мозга, и удаление больших полушарий практически их не изменяет. У кошек после перерезки ствола между средним и промежуточным мозгом локомоция сохраняется лишь частично. Выключение коры большого мозга в эксперименте у обезьян и в патологических случаях у человека приводит к потере не только произвольных движений, но и выпрямительных рефлексов. В процессе обработки информации в нейронах коры большого мозга различные отделы коры играют специфическую роль.
Поиск по сайту: |