|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Глава 6.1. Элементный составСтр 1 из 4Следующая ⇒
ЧАСТЬ 6. ХИМИЧЕСКИЙ СОСТАВ ЖИВЫХ ОРГАНИЗМОВ. ПОТРЕБНОСТИ В ВЕЩЕСТВАХ И ЭНЕРГИИ Глава 6.1. Элементный состав
6.1.1. Классификации элементов, входящих в состав живых организмов. По отношению элементов к живым организмам принято разделение их на несколько групп: 1) на биогенные и абиогенные, 2) на макро-, микро- и ультрамикроэлементы, 3) на ятрогенные (т. е. вроде бы и нужные, но вредящие), эссенциальные (жизненно важные) и условно эссенциальные, 4) на токсичные и условно токсичные. Из 92 стабильных в естественных условиях элементов в живых организмах обнаружен 81. В табл. 1 показано содержание некоторых химических элементов в растительных и животных организмах, приведенное в перерасчете на количество молей на тонну сухого органического вещества. Биогенные элементы − это химические элементы, постоянно входящие в состав организмов и имеющие определённое биологическое значение. К ним прежде всего следует отнести кислород (составляющий 70 % массы организмов), углерод (18 %), водород (10 %), кальций, азот, калий, фосфор, магний, сера, хлор, натрий, железо. Эти элементы входят в состав всех живых организмов, составляют их основную массу и играют большую роль в процессах жизнедеятельности. Многие элементы имеют большое значение только для определённых групп живых существ (например, бор необходим для растений, ванадий — для асцидий). Содержание тех или иных элементов в организмах зависит не только от их видовых особенностей, но и от состава среды, пищи (для растений − от концентрации и растворимости тех или иных почвенных солей), экологических особенностей организма и других факторов. Биогенные элементы, в свою очередь, подразделяются на макро-, микро- и ультрамикроэлементы (табл. 2). Первую подгруппу биогенных элементов образуют первоэлементы, или органогены. Исходя из весовых отношений (т. е. г/т), к органогенам традиционно причисляют, учитывая их общее содержание
Таблица 1 Содержание некоторых химических элементов в раститель-
в живом веществе (98,72 ат %), четыре элемента: кислород, углерод, водород и азот. Эти четыре элемента идеально подходят к выполнению биологических функций, так как все они легко образуют ковалентные связи посредством спаривания электронов. Для того чтобы полностью укомплектовать свои внешние электронные оболочки и образовать таким образом стабильные ковалентные связи, водороду требуется один электрон, кислороду – два, азоту – три и углероду – четыре электрона. Эти четыре элемента могут легко реагировать друг с другом, заполняя свои внешние электронные оболочки. С помощью одинарных и двойных связей углерод, азот и кислород могут образовывать различные химические соединения. Среди элементов, способных образовывать ковалентные связи, они самые легкие. Так как прочность ковалентной связи обратно пропорциональна атомным весам связанных с ее помощью атомов, то возможно, что живые организмы «выбрали» именно эти элементы из-за их способности формировать прочные ковалентные связи. Очень важна способность атомов углерода взаимодействовать друг с другом, образуя стабильные углерод-углеродные связи, что и обеспечивает углеводородные каркасы разнообразных молекул. Соединениям углерода свойственна еще одна отличительная особенность, которая состоит в способности спаренных электронов образовывать вокруг каждого атома углерода тетраэдрическую конфигурацию, благодаря чему различные типы органических молекул обладают разной трехмерной структурой. Никакой другой химический элемент, кроме углерода, не может создавать стабильные молекулы со столь разнообразными конфигурациями и размерами и с таким многообразием функциональных групп. Кремний, значительно более распространенный в литосфере, чем углерод, оказался менее пригодным для живых организмов, по всей видимости, потому, что: 1) соединения кремния малорастворимы; 2) в присутствии кислорода связи кремний – кремний нестабильны. В качестве первичных элементов жизни необходимо добавить еще фосфор и серу. В самом деле, жизнь невозможна без передачи наследственной информации и без энергии. Фосфор входит в состав основы молекул наследственности – ДНК и РНК, а также молекулы, поставляющей энергию, – АТР. Сера не только входит в состав двух из 20 основных аминокислот – метионина и цистеина − единиц, образующих белки всех высших организмов, но и в их составе, образуя дисульфидные мостики, формирует третичную, глобулярную структуру белка. Во вторую подгруппу биогенных элементов под общим названием «макроэлементы» можно объединить шесть элементов: калий, натрий, кальций, магний, хлор, кремний. Эти элементы явились основой для появления одноклеточных организмов около 3 млрд лет назад. Необходимо было развить системы внутреннего минерального гомеостаза посредством создания клеточной мембраны и тургорной, буферной системы. Третья подгруппа биогенных элементов – эссенциальные микроэлементы, удовлетворяющие следующему условию: и дефицит, и избыток данного элемента приводят к патологическим отклонениям в организме. Это направление было развито с использованием лабораторных животных, содержащихся на искусственных диетах, обедненных или обогащенных тем или иным микроэлементом. Кроме того, необходимые знания были получены при изучении заболеваний различной этиологии у человека. К этой подгруппе можно отнести десять элементов, биологическая значимость которых в организме высших млекопитающих и человека на сегодняшний день твердо установлена: железо, медь, цинк, марганец, хром, селен, молибден, йод, кобальт, фтор. Существует ряд микроэлементов, стабильно присутствующих в человеческом организме. Дефицитные их состояния обнаружены лишь у некоторых сельскохозяйственных и лабораторных животных. В четвертую подгруппу условно эссенциальных микроэлементов относятся следующие: мышьяк, бор, бром, литий, никель, ванадий, кадмий, свинец. В геологическом плане большинство из них вулканического происхождения, появились на относительно поздних этапах развития Земли, и можно предполагать, что в метаболизм организмов с эволюционной точки зрения они включились сравнительно поздно. Имеется группа элементов с неизведанными функциями, которую с достаточно большой степенью осторожности можно связать с интеллектуальными способностями человека. Так, обращает на себя внимание относительно высокое содержание в головном мозге золота (2,54 мкмоль/кг сухой массы), таллия (2,44 мкмоль/кг, тогда как в других органах – не более 1,96 мкмоль/кг), олова (16 мкмоль/кг, что на порядок превышает его содержание в других органах) и некоторых других элементов. При исследовании детей одного из районов Новосибирской области с диагнозом «умственная отсталость» было установлено, что у больных детей в волосах достоверно повышена концентрация марганца, ванадия и никеля и понижена концентрация галлия. Все эти элементы можно объединить в пятую подгруппу брэйн-элементов. Понятие «токсичные элементы» не имеет права на существование, так как избыток любого химического элемента в человеческом организме приводит к патологии. Это касается элементов всех групп, даже первоэлементов. Печально известная синильная кислота имеет формулу HCN, т. е. представляет собой соединение трех органогенов. Следовательно, речь можно вести только о токсичной концентрации либо о токсичных соединениях того или иного элемента. Кроме биогенных элементов, существуют абиогенные, которые подразделяются на три подгруппы (см. табл. 2). К первой можно отнести элементы-нейтралы типа алюминия, титана и рубидия, относительно высокие концентрации которых организм переносит достаточно безболезненно. Поскольку на ранних этапах эволюции органического мира организмы имели возможность столкнуться с их высокой концентрацией во внешней среде (древнем океане и литосфере), в течение сотен миллионов лет они выработали механизмы толерантности. Эволюционно более «молодые» элементы, имеющие в основном вулканическое происхождение, а также появившиеся в самые последние годы техногенные (ртуть, висмут, осмий и т. д.), к высоким концентрациям которых во внешней среде организмы не в состоянии были приспособиться на протяжении жизни ограниченного числа поколений, относят ко второй группе агрессивных абиогенных элементов. Существует третья подгруппа элементов-конкурентов, представленных в древнем океане: барий, стронций и др. Составляя скелетную основу некоторых форм жизни, а после выхода организмов Таблица 2 Биогенная классификация химических элементов [1]
на сушу и замены этих элементов более легким и реакционноспособным кальцием, они стали конкурировать с ним за место в метаболизме организмов. Например, болезнь, возникающая при замещении кальция на стронций в скелетах людей в некоторых эндемичных районах Сибири, вызывает хрупкость кости и недоразвитие скелета. Таким образом, абиогенные элементы можно разделить на три подгруппы: агрессивные, нейтральные и элементы-конкуренты. Из всего сказанного можно сделать вывод: – первоэлементы являются сквозными для всех форм жизни на Земле, т. е. присущи всем формам жизни; – макроэлементы – сквозные для всех животных организмов; – эссенциальные микроэлементы – сквозные для всех млекопитающих; – условно эссенциальные – сквозные для отдельных семейств млекопитающих; – брэйн-элементы – сквозные для высших млекопитающих и человека. Перейдем к рассмотрению отдельных элементов, входящих в состав живых организмов. 6.1.2. Кислород.Кислород необходим организму для протекания многочисленных реакций окисления, в том числе для функционирования цепи переноса электронов (ЦПЭ), которая потребляет до 90 % поступающего в клетки кислорода. 6.1.2.1. Активные формы кислорода.Во многих клетках организма в результате последовательного (поэтапного) одноэлектронного присоединения четырех электронов к одной молекуле кислорода образуются так называемые активные формы кислорода, к которым относятся супероксидный радикал, пероксид водорода и гидроксильный радикал:
е е, 2Н+ е, Н+ О2 → О2− → Н2О2 → Н2О + ОН• (1) Конечным продуктом этих реакций является вода. Из схемы (1) видно, что образуются два радикала, наиболее активный из них − гидроксильный радикал, вступающий в реакции со многими органическими молекулами. В невозбужденном состоянии кислород нетоксичен. Молекула кислорода содержит два неспаренных электрона с параллельными спинами. Они не могут образовывать термодинамически стабильную пару и располагаются на разных орбиталях, каждая из которых способна принять по одному электрону. Образовавшиеся радикалы способны отнимать электроны от многих органических соединений и инициировать цепные реакции окисления, приводящие к нарушению структуры и функций биополимеров. Так, в белках происходит окисление некоторых аминокислот (например, тирозина), образование поперечных сшивок, наблюдается накопление гликированных белков (см. ч. 5, с. 142−148) − все это приводит к изменению третичной структуры белковой молекулы и потере ею функциональной активности. Кроме этого, перечисленные выше реакции приводят к активации протеолитических ферментов, гидролизующих модифицированные белки. В нуклеиновых кислотах активные формы кислорода вызывают окислительную модификацию гетероциклических оснований с последующим гидролизом гликозидной связи между модифицированным основанием и остатком сахара и разрывами в двуцепочечной структуре, что в конечном счете может привести к нарушению целостности хромосом, а также к активации нуклеаз, расщепляющих поврежденные участки нуклеиновых кислот. Свободнорадикальное окисление затрагивает и клеточные липиды, приводя к нарушению мембранной структуры клетки, что может закончиться ее гибелью. 6.1.2.2. Пути образования активных форм кислорода. Основным путем образования активных форм кислорода является утечка электронов из ЦПЭ (при функционировании QH2-дегидрогеназного комплекса, принимающего от донора последовательно по одному электрону с образованием промежуточной формы кофактора − семихинона СоQ•) и взаимодействие промежуточных радикальных форм с молекулой кислорода (в случае семихинона СоQ• с образованием супероксидного радикала). В случае цитохромоксидазы (комплекс IV) «утечки» электронов не наблюдается, так как кислород восстанавливается в специальных активных центрах фермента, содержащих ионы железа и меди, без высвобождения промежуточных свободных радикалов из активного центра. Активные формы кислорода образуются также в реакциях, катализируемых оксидазами и оксигеназами. Оксидазы (оксидазы аминокислот, супероксиддисмутаза, пероксисомальные оксидазы) восстанавливают кислород с образованием пероксида водорода:
SH2 + O2 → S + H2O2 ,
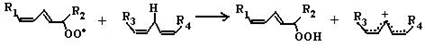
где SH2 − окисляемый субстрат. Эти ферменты используют кислород лишь в качестве акцептора электронов, восстанавливая его до воды или пероксида водорода. Источниками активных форм кислорода могут быть также монооксигеназы (например, цитохром Р450), включающие один атом кислорода в продукт реакции, и диоксигеназы, включающие оба атома кислорода в окисляемую молекулу. В присутствии Fe2+ или ионов других переходных металлов пероксид водорода способствует образованию наиболее токсичной формы кислорода − гидроксильного радикала (ОН•): Fe2+ + H2O2 → Н2О + ОН• (2) В лейкоцитах в процессе фагоцитоза усиливается образование активных форм кислорода, катализируемое NADPH-оксидазой. Этот фермент локализован преимущественно на наружной стороне плазматической мембраны, инициируя так называемый «респираторный взрыв» с образованием активных форм кислорода. 6.1.2.3. Перекисное окисление липидов (ПОЛ). Действию активных форм кислорода особенно подвержены ненасыщенные жирные кислоты, содержащие двойные связи, расположенные через метиленовую (-СН2-)-группу. Отнимая электрон от метиленовой группы, радикальные формы кислорода (чаще всего гидроксильный радикал), являющиеся инициаторами окисления липидов, превращают липид, содержащий эту кислоту, в липидный радикал. ПОЛ представляет собой цепные реакции, обеспечивающие воспроизводство частиц, содержащих неспаренный электрон, и инициирующие дальнейшее распространение перекисного окисления. 1. Инициация ПОЛ: образование свободного липидного радикала (L•): -СН2- → -СН•-(3)
L → L• 2. Развитие цепи: L• + О2 → LOO•(4)
LOO• + LH → LOOH + L•(5)
Развитие цепи путем присоединения молекулы кислорода приводит к образованию пероксирадикала LOO• или пероксида липида LOOH. Таким образом, каждый образовавшийся радикал инициирует образование несколько других. 3. Обрыв цепи путем взаимодействия радикалов между собой: LOO• + L• → LOOH + LH(6)
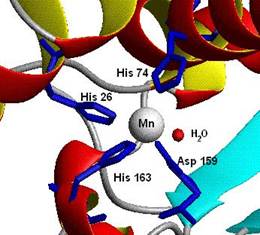
L• + витамин Е → LH + витамин Е•(8) витамин Е• + L• → LH + витамин Еокисл.(9) Развитие цепи обрывается в результате взаимодействия свободных радикалов между собой или взаимодействия с антиоксидантами, например с витамином Е, который превращается в стабильную окисленную форму при отдаче электронов. Конечными продуктами ПОЛ являются малоновый диальдегид и гидропероксид полиеновой кислоты. 6.1.2.4. Нарушения в клетке, вызываемые ПОЛ. Из-за образования гидропероксидов полиеновых кислот в составе липидов в гидрофобных клеточных мембранах появляются гидрофильные зоны, через которые в клетку проникает вода, катионы металлов, что приводит к разбуханию клетки и ее разрушению. ПОЛ наблюдается при многих заболеваниях: при дистрофии мышц, болезни Паркинсона, атеросклерозе, развитии опухолей. ПОЛ имеет место также при спазме коронарных артерий (ишемии) и последующем их расширении при реоксигенации. На коже изменение структуры тканей в результате ПОЛ наблюдается в виде увеличивающегося с возрастом количества пигментных пятен. Прогоркание жиров при хранении, появление неприятного запаха у молочных продуктов также являются результатом ПОЛ. 6.1.2.5. Системы защиты клетки от активных форм кислорода.Для защиты от активных форм кислорода в организме существуют эффективные системы ингибирования их действия. К этим системам относятся как ферменты антиоксидантного действия (супероксиддисмутаза, каталаза, глутатионпероксидаза), так и витамины-антиоксиданты (витамин Е, витамин А и каротиноиды, аскорбиновая кислота, биофлавоноиды). В органах, где много митохондрий, цитохрома Р450 и пероксисом (печени, надпочечниках и почках), ферменты антиоксидантного действия наиболее активны. Супероксиддисмутаза Супероксиддисмутаза (К.Ф. 1.15.1.1) катализирует превращение супероксидного радикала, который образуется первым из активных форм кислорода при утечке электронов из дыхательной цепи, в пероксид водорода: 2 О2− + 2 Н+ → Н2О2 + О2 (10) Изоферменты супероксиддисмутазы обнаружены в цитоплазме, в митохондриях клеток, а также во внеклеточном пространстве. Это индуцируемый фермент, количество которого увеличивается при активации перекисного окисления. Превращение супероксида, катализируемое супероксиддисмутазой, можно разбить на две реакции: M(n+1)+ − СОД + O2− → Mn+ − СОД + O2, Mn+ − СОД + O2− + 2H+ → M(n+1)+ − СОД + H2O2, где M представляет собой переходный металл, а именно Cu (n=1); Mn (n=2); Fe (n=2); Ni (n=2). В данной реакции окислительное состояние катиона металла изменяется между n и n + 1. Существует несколько форм супероксиддисмутазы в зависимости от типа переходного металла, играющего роль кофактора активного центра фермента: Cu, Zn-СОД, в которой медь выступает кофактором активного центра, а цинк стабилизирует конформацию, Mn-СОД (с марганцем в активном центре), а также менее распространённые Fe-СОД (с железом) и Ni-СОД (с никелем). Cu, Zn-СОД − самая распространённая супероксиддисмутаза, обнаруженная в цитозоле практически всех эукариотических клеток, является гомодимером молекулярной массой 32,5 kDа. Cтруктура гомодимера стабилизирована образованием гидрофобных и элект-ростатических связей. Медь и цинк связаны с белковой частью молекулы через гистидиновые остатки. Митохондрии эукариотических клеток и многие бактерии содержат Mn-СОД, представляющую собой гомотетрамер, содержащий по одному иону марганца на каждую субъединицу, например митохондриальная СОД человека (К.Ф. 1.15.1.1) молекулярной массой 86−88 kDа. Марганец в этом ферменте координирован с белком через три гистидина и аспартат, а также с водой либо гидроксильным лигандом в зависимости от окислительного состояния марганца (Mn2+ или Mn3+) (рис. 6.1).
Рис. 6.1. Структура активного центра митохондриальной Mn-супероксиддисмутазы человека (согласно [4]) E. coli и многие другие бактерии содержат формы фермента с железом в активном центре (Fe-СОД). У бактерий Mn-СОД и Fe-СОД представлены как гомодимерными, так и гомотетрамерными формами. СОД является одним из наиболее термостабильных глобулярных белков: выдерживает нагревание до 80 оС, сохраняет свою активность даже в 8 М растворе мочевины. Супероксиддисмутаза, поддерживая стационарную концентрацию супероксидных радикалов на определенном уровне, защищает тем самым клеточные структуры от повреждающего действия как самих радикалов О2•-, так и от появления гидроксильных радикалов, которые могут образовываться из супероксидного радикала. СОД играет важную роль в процессе старения организмов и при некоторых заболеваниях, таких как гемолитическая анемия, ишемия, ряд нейрологических заболеваний. Существенная роль в развитии воспалительных процессов отводится супероксидным радикалам. Успешное лечение воспалительных процессов с помощью СОД, например при артритах, позволяет рассматривать этот фермент как альтернативу кортикостероидам. Каталаза Образовавшийся пероксид водорода подвергается превращениям под действием каталазы: 2 Н2О2 → 2 Н2О + О2(11) Каталаза (К.Ф. 1.11.1.6) локализована в основном в пероксисомах, где образуются наибольшие количества пероксида водорода, и в лейкоцитах для защиты клеток от последствий «респираторного взрыва» (см. п. 6.1.2.2). Каталаза в зависимости от структуры относится к одному из трех семейств. Семейство 1 каталаз − это тетрамерные белки, состоящие из больших или малых субъединиц, содержащих гем. Гем включается в фермент во время его сборки в пероксисомах из мономеров, которые гем не содержат. Семейство 2 − это бифункциональные ферменты, проявляющие и каталазную, и пероксидазную активности и являющиеся гемсодержащими асимметричными димерами. Семейство 3 − гексамерные белки, содержащие по два иона марганца на мономер (рис. 6.2). Реакция, катализируемая семействами 1 и 2 каталаз, состоит из двух стадий: сначала один атом кислорода из пероксида водорода включается в состав молекулы воды, а второй восстанавливается гемом с образованием оксоферрильного порфирин-π-радикального комплекса (compound I) (один электрон передается ионом железа, второй происходит из порфиринового кольца) (см. схему (12)). Во время второй стадии реакции оксоферрильное производное взаимодействует со второй молекулой пероксида водорода с образованием молекул воды, кислорода и восстановлением исходной формы фермента (схема (13)). Enz (Por-Fe3+) + H2O2 → Cpd I (Por+.-Fe4+ = O) + H2O(12) Cpd I (Por+.-Fe4+= O) + H2O2 → Enz (Por-Fe3+) + H2O + O2 ,(13) где Enz − каталаза, Por− порфирин, Cpd I− промежуточное производное I (compound I).
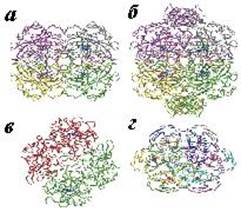
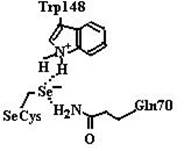
Рис. 6.2. Пространственная структура каталазы: а − тетрамер, состоящий из малых субъединиц (Pseudomonas syringae); б − тетрамер, в состав которого входят большие субъединицы (E. сoli); в − димерная структура бифункционального фермента каталазы-пероксидазы (Burkholderia pseudomallei); г − гексамер Мn2-содержащей каталазы (Lactobacillus plantarum). Гем в структурах а и б изображен синим цветом, ионы Мn в структуре г − красным (согласно [14]) В случае пероксидазной реакции, катализируемой ферментами семейства 2, необходим органический донор электронов для протекания второй стадии (8): Cpd I (Por+.-Fe4+= O) + 2AH2 → Enz (Por-Fe3+) + 2AH• + H2O,(14) где AH2 является органическим соединением − донором электронов. Особенностью этой группы ферментов является наличие в структуре ковалентного аддукта Мet-Tyr-Trp, в случае его отсутствия протекает только пероксидазная реакция. Каталазы, относящиеся к семейству 3, катализируют реакции превращения пероксида водорода, где роль восстановителя выполняет комплекс из двух катионов марганца: H2O2 + Mn2+-Mn2+ (2H+) →Mn3+-Mn3+ + 2 H2O(15) H2O2 + Mn3+-Mn3+ →Mn2+-Mn2+ (2H+) + O2, (16) т. е. центр, содержащий два иона марганца, ведет себя аналогично гему в каталазах семейств 1 и 2. Глутатионпероксидаза Огромное значение для инактивации активных форм кислорода имеет глутатионпероксидаза (К.Ф. 1.11.1.9), катализирующая восстановление пероксида водорода и липидных гидропероксидов с помощью глутатиона − γ-глутамилцистеинилглицина (GSH). Сульфгидрильная группа GSH окисляется до дисульфидной формы, отдавая электроны пероксиду водорода или гидропероксиду липида: 2 Н2О2 + GSH → 2 Н2О + G-S−S-G.(17) Окисленный глутатион восстанавливается под действием глутатионредуктазы (К.Ф. 1.6.4.2): G-S−S-G + NADPH + H+ → 2 GSH + NADP+.(18) В организме животных в зависимости от органа и ткани обнаружено несколько форм глутатионпероксидаз. Глутатионпероксидаза чаще всего представляет собой гомотетрамер. В активном центре большинства из них присутствует остаток селеноцистеина, необходимого для протекания ферментативной реакции. На рис. 6.3 приведена схема реакции, катализируемой глутатионпероксидазой. На первой стадии реакции остаток селеноцистеина, ионизованная форма которого стабилизируется остатками Trp148 и Gln70 (нумерация аминокислотных остатков приведена для фермента из бычьих эритроцитов, но оба остатка, являясь консервативными, присутствуют практически во всех глутатионпероксидазах), окисляется под действием пероксидов до селенильной кислоты RSeOH. Следующие две стадии реакции заключаются в регенерации селеноцистеина с помощью двух молекул глутатиона с образованием промежуточного продукта селененилсульфида (EnzSeSG).
Рис. 6.3. Схема реакции, катализируемой глутатионпероксидазой. Ионизованная форма остатка селеноцистеина в активном центре фермента стабилизируется остатками Trp148 и Gln70 (схема справа). Под действием пероксидов происходит окисление селеноцистеина до селенильной кислоты (схема слева), которая восстанавливается обратно в селеноцистеин двумя молекулами GSH с образованием промежуточного соединения селенилсульфида (EnzSeSG) (согласно [23]) Таким образом, недостаток селена в организме приводит к увеличению активных форм кислорода в клетках, что, в свою очередь, проявляется в ослаблении иммунной системы организма и возникновению заболеваний, связанных с многочисленными изменениями в структуре и функциях биополимеров (белков, нуклеиновых кислот и липидов). Восстановление дисульфидной формы глутатиона (G-S−S-G) происходит в организме с помощью флавопротеина глутатионредуктазы, представляющей собой гомодимер и состоящей из нескольких доменов: FAD- и NADPH-связывающих, домена для взаимодействия мономеров при образовании димера. FAD-связывающий домен располагается на N-конце молекулы фермента, как видно из рис. 6.4. Восстановление димерной формы глутатиона происходит за счет передачи электронов от NADPH (NADH) через FADH2 к G-S−S-G (схема (12)).
Рис. 6.4. Пространственная структура гомодимерных белков, входящих в семейство глутатионредуктазы. Домены фермента выделены цветом: FAD-связывающий − желтым, NADP-связывающий − бирюзовым, домен, связывающий мономеры друг с другом, − оранжевым (согласно [32]) Витамин Е Один из наиболее распространенных природных антиоксидантов витамин Е (структура соединений, входящих в семейство витамина Е, приведена на схеме (19)), являясь липофильной молекулой, взаимодействует с активными формами кислорода непосредственно в гидрофобном слое мембран, предотвращая развитие цепи ПОЛ.
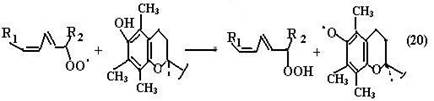
Витамин Е восстанавливает свободный радикал пероксида липида (LOO•) до гидропероксида LOOH, тем самым останавливая ПОЛ.
Витамин С Аскорбиновая кислота (витамин С) (структура и превращения аскорбиновой кислоты в результате окислительно-восстановитель- ной реакции приведены на схеме (21)) может принимать участие в ингибировании реакций ПОЛ по двум направлениям: 1) она восстанавливает окисленную форму витамина Е, тем самым поддерживая необходимую его концентрацию в мембранах клеток; 2) витамин С восстанавливает водорастворимые активные формы кислорода (супероксидный радикал, пероксид водорода и гидроксильный радикал), инактивируя их.
Витамин А Витамин А (ретинол) и его предшественники − каротиноиды (α-, β-каротины), структура которых приведена на схеме (22), обладая антиоксидантным действием, также активно ингибируют развитие ПОЛ.
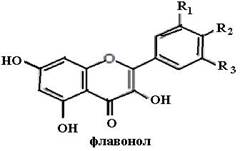
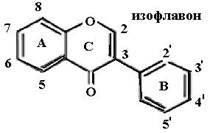
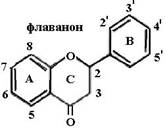
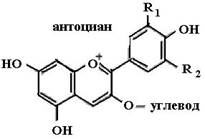
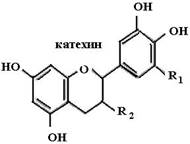
Биофлавоноиды К системам защиты организма от действия активных форм кислорода относят также биофлавоноиды. Эти соединения являются полифенолами. Среди них различают флавоны, флавонолы, флаваноны, антоцианы, катехины (рис. 6.5), обнаруженные в больших количествах в составе растений, например, в чайном листе. К наиболее
Рис. 6.5. Структура биофлавоноидов, среди которых различают флавонолы, флавоны, изофлавоны, флаваноны, антоцианы, катехины известным и используемым среди биофлавоноидов относят кверцетин и рутин:
Механизм защитного действия биофлавоноидов еще недостаточно выяснен. Известно, что переходные металлы (железо) в организме реагируют с супероксидным радикалом с образованием пероксида водорода: [2Fe2+2Fe3+−4S] + O2•− + 2H+ → [Fe2+3Fe3+−4S] + H2O2.(24) Пероксид водорода в присутствии переходного металла образует гидроксильный радикал, наиболее активно модифицирующий клеточные биополимеры: Fe2+илиCu+ + H2O2 + H+ → Fe3+илиCu2+ + •OH + H2O(25)
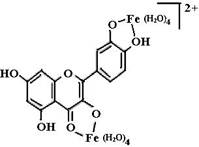
Предполагают, что биофлавоноиды, являясь полифенолами, способны связывать переходные металлы (железо, медь), тем самым уменьшая образование гидроксильных радикалов (схема (27)) и защищая организм от их действия.
На схеме (28) представлены комплексы железа с рутином и кверцетином.
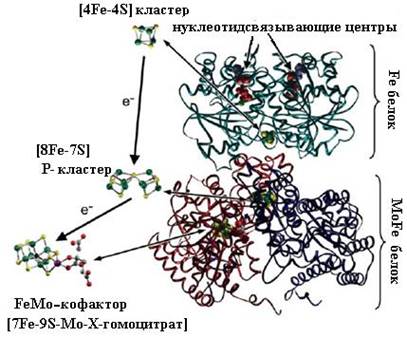
6.1.3. Азот.В окружающей среде азот встречается в виде молекулярного азота, неорганических (нитриты, нитраты, аммонийные соли и т. д.) и органических (мочевина, амины, аминокислоты, нуклеотиды) соединений. Значение органических соединений азота для организмов велико. Стоит только упомянуть, что ни один живой организм не способен существовать без белков и нуклеиновых кислот, а также их мономеров − аминокислот и нуклеотидов. Так для синтеза аминокислот необходим аммиак. Большинство организмов получают его из пищи путем восстановления нитратов или нитритов или гидролиза потребляемых белков. Атомы в молекуле азота связаны прочной тройной ковалентной связью, из-за чего он практически не вступает в реакции окисления-восстановления в нормальных условиях без применения катализаторов и не может использоваться растениями и животными. Лишь небольшая часть организмов способна синтезировать аммиак из молекулярного азота. 6.1.3.1. Азотфиксация. Для получения азотных удобрений, необходимых для развития культивируемых растений, человечество ежегодно затрачивает огромные деньги, при этом в больших количествах образуются токсические отходы, загрязняющие окружающую среду. Микроорганизмы в течение миллионов лет незаметно и чисто восстанавливают молекулярный азот до иона аммония, затрачивая на это энергию гидролиза АТР и восстановительные эквиваленты NADH, а также используя серию ферментов (ферредоксин, гидрогеназа), важнейшим из которых является нитрогеназа (К.Ф. 1.18.6.1). Этот процесс называется азотфиксацией и осуществляется азотфиксирующими микроорганизмами, в наземных экосистемах обитающих в основном в почве (это представители трех семейств: Azotobacter, Clostridium, Rhizobium). Некоторые из них, например клубеньковые бактерии (сем. Rhizobiaceae), существуют в симбиозе с бобовыми растениями. Азотфиксаторы живут также в кишечнике многих животных (жвачные, грызуны, термиты) и человека (род Escherichia). Восстановление азота состоит из трех стадий, на каждой из которых происходит присоединение к восстанавливаемой молекуле пары электронов и двух протонов: N2 + 2 e + 2 H+ → NH=NH + 2 e + 2 H+ → NH2−NH2 + 2 e + + 2 H+ → 2 NH3 (29) Общее уравнение процесса азотфиксации можно записать в следующем виде: N2 + 8 e + 8 H+ + 16 MgATP → 2 NH3 + 16 MgADP + + 16 Pi + H2 (30) Нитрогеназа представляет собой комплекс из двух ферментов: собственно нитрогеназа, содержащая Mo-Fe-белковый кофактор и называемая также динитрогеназой, и редуктаза нитрогеназы (Fe-белок). Оба фермента сохраняют свои каталитические свойства только в комплексе, при разделении они инактивируются. Ферментативной активностью обладают также «гибриды», например, из Mo-Fe-белкового кофактора азотобактерии с Fe-белком клубеньковых бактерий. Mo-Fe-Белковый кофактор (молекулярной массой 200−250 kDa) состоит из четырех субъединиц двух типов (α и β), содержит 2 атома Мо (в виде FeMo-кофактора), 28−34 атома негемового железа и 18−24 атома сульфидной серы. Fe-Белок (молекулярной массой 50−60 kDa) состоит из двух одинаковых субъединиц. Для своего функционирования нитрогеназа нуждается в непрерывном притоке как энергии (в форме АТР), так и электронов. Это обеспечивается благодаря дыханию и брожению, происходящим в микроорганизмах, или в результате фотосинтеза (в цианобактериях). АТР специфически связывается с Fe-белком (две молекулы на димер), и образовавшийся комплекс переносит электроны от ферредоксина или флаводоксина с помощью железо-серного кластера [4Fe−4S] (одного на димерную молекулу Fe-белка) к Mo-Fe-белковому кофактору.
После переноса восстановительных эквивалентов Fe-белок диссоциирует из комплекса, а восстановленный Mo-Fe-белок связывает N2 в кластере, названном FeMo-кофактор [7Fe–9S–Mo–X−гомоцит- рат] (Х по расчетам может быть атомом углерода, азота или кислорода) и локализованном на α-субъединицах тетрамера, и последовательно восстанавливает молекулярный азот до NH3. На рис. 6.6 представлена пространственная структура нитрогеназы Azotobacter vinelandii.
Рис. 6.6. Пространственная структура нитрогеназы Azotobacter vinelandii. Зелёным показан Fe-белок, синим и фиолетовым — MoFe-белок. Fe-белок содержит сайт связывания нуклеотида (АТР) и [4Fe-4S] кластер для переноса восстановительных эквивалентов, MoFe-белок через Р-кластер [8Fe-7S], соединяющий α- и β-субъединицы в димерах, передает электроны на FeMo-кофактор [7Fe-9S-Mo-X-гомоцитрат], локализованный на α-субъединицах. На рисунке изображена половина нитрогеназного комплекса: один Fe-белок и αβ-структура из α2β2-тетрамера MoFe-белкового кофактора (согласно [10]) Каталитический центр Mo-Fe-белкового кофактора основан на [4Fe-3S]-кластере, связанном тремя сульфидными связями с [Mo-3Fe-3S]-субкластером, гомоцитрат координирован с ионом молибдена через два атома кислорода (схема (32)) (положение аминокислотных остатков приведено для нитрогеназы Klebsiella pneumoniae). Молекула азота связывается четырьмя ионами железа.
Fe-белок не только переносит восстановительные эквиваленты от ферридоксина (флаводоксина) к FeMo-кофактору, но и необходим при сборке Mo-Fe-белкового кофактора для встраивания комплекса Мо-гомоцитрат в предшественник кофактора, не содержащий иона молибдена. В его отсутствие активный Mo-Fe-белковый кофактор не образуется. Нитрогеназа обладает низкой субстратной специфичностью и вместо азота может восстанавливать другие соединения с тройной связью: ацетилен (на этом её свойстве основан метод определения активности азотфиксации), цианиды и азиды (также параллельно с образованием водорода). Как уже было сказано, что восстановление молекулы азота происходит последовательно. На рис. 6.7 представлен цикл азотфиксации. Видно, что сначала восстанавливается один атом азота, потом другой, что отличается от реакций, приведенных на схеме (29). Пока неясно, какая из схем более правильно описывает процесс восстановления молекулярного азота до иона аммония. В 1980−1990-е годы у ряда бактерий (Azotobacter chroococcum, Az. vinelandii, Az. paspalii, Anabaena variabilis, Rhodobacter capsulatus) обнаружены формы нитрогеназы, содержащие вместо иона молибдена ионы ванадия, вольфрама или железа, они встраиваются в фермент при дефиците молибдена. Эти ферменты устроены аналогично нитрогеназе, содержащей ион молибдена. Их уникальным свойством является способность восстанавливать ацетилен в этан, в то время как Мо-нитрогеназа восстанавливает ацетилен в этилен.
Рис. 6.7. Цикл восстановления молекулярного азота нитрогеназой (согласно [10]) В 1997 г. у термофильных бактерий (Streptomyces thermoautotrophicus) была открыта супероксидзависимая (СОЗ) нитрогеназа. Проблемой функционирования всех металлсодержащих нитрогеназ является инактивация при контакте с кислородом. Организмы создали разнообразные системы защиты от инактивации нитрогеназы кислородом: 1) разделение во времени процессов фотосинтеза и азотфиксации, например процесс азотфиксации протекает ночью в отсутствие освещения, 2) создание дифференцированных клеток-гетероцист с анаэробными или микроаэробными условиями. СОЗ-нитрогеназа в отличие от других нитрогеназ не только не инактивируется в атмосфере кислорода, но и требует его присутствия для проявления активности. Другое ее отличие − она трехкомпонентна. Хотя активность «альтернативных» нитрогеназ составляет не более 10 % от активности Мо-нитрогеназы, наличие этих ферментов позволяет азотфиксаторам вести процесс при низком содержании молибдена, что характерно для многих типов почв. Часть энергии, используемой для азотфиксации, теряется организмами, так как не все из них способны использовать водород, образующийся в реакции (схема (30)). Образовавшийся аммиак глутаминсинтетазой включается в состав глутамата: Glu + NH3 + ATP → Gln + ADP + Pi (33) и в виде глутамина доставляется ко всем клеткам организма, где используется для синтеза азотсодержащих органических соединений. 6.1.3.2. Кислородсодержащие соединения азота.В живых организмах обнаружено большое количество кислородсодержащих неорганических и органических соединений азота. К ним относятся окислы азота, ион нитрозония, пероксинитрит, нитриты, нитраты и органические нитросоединения. Окись азота NO. Является одним из важных соединений, участвующих в межклеточной сигнализации. Ее синтез и функции будут подробно рассмотрены в ч.7. Здесь мы кратко рассмотрим свойства данного соединения. NO представляет собой гидрофобный газ. Он может существовать в виде относительно стабильного, нейтрально заряженного радикала (NO•) с липофильными свойствами и выраженной тенденцией взаимодействовать с молекулами, обладающими неспаренным электроном, такими как супероксидный анион, железо и молекулярный кислород. NO• может также подвергаться одноэлектронному восстановлению с образованием нитроксил аниона (NO-) или, потеряв электрон, превращается в ион нитрозония (NO+). Ион нитрозония NO+. Легко реагирует с SH-группами, образуя биологически активные S-нитрозосоединения. Очевидно, именно механизм нитрозилирования сократительных регуляторных белков и белков ионных каналов лежит в основе расслабления гладких мышц при их спазме. Пероксинитрит. Превращение NO. из физиологического регулятора в токсический агент происходит в результате взаимодействия NO c супероксидным анионом с образованием аниона пероксинитрита ONOO- (схема (34)), который, распадаясь в процессе диффузии от места его образования на гидроксильный радикал ОН- и двуокись азота NO2, сокрушает все на своем пути: биополимеры и биомембраны. O2•− + NO• → ONOO−(34) Аниона пероксинитрита в нормальных условиях образуется мало. Если активность NO-синтаз низкая, то образовавшийся NO. легко нейтрализуется кислородом. В случае же массового производства NO. и супероксидного аниона равновесие сдвигается в сторону образования аниона пероксинитрита, и тогда наступает катастрофа для клеточного окружения. Сам ONOO- и продукты распада его протонированной формы повреждают или разрушают биологические структуры путем их окисления или нитрозилирования. Распад аниона пероксинитрита происходит тремя разными способами. Поскольку анион ипероксинитрит имеет рКа, равную 6,9, то при физиологических значениях рН он подвергается протонированию: ONOO- + Н+ → ONOOН. (35) Затем ONOOН или распадается на гидроксильный радикал и двуокись азота после взаимодействия с клеткой-мишенью: ONOOН → ОН● + NO2, (36) или в отсутствие «жертвы» распадается до нитратов: ONOOН → HNO3 ↔ NO3− + H+. (37) Третий путь распада аниона пероксинитрита наиболее сложен: ONOO− + металл → NO+2 ...O− - - - металл → белок - - тирозин −NO2 + HO-металл1+ + металл 2+ + Н2О (38) Следствием этой реакции in vivo является модификация (нитрование) тирозиновых остатков белковых молекул. Это приводит к деструкции многих ферментных и структурных систем и блокированию клеточной сигнализации, опосредуемой тирозинкиназой, так как образующийся в ходе реакции нитротирозин очень напоминает фосфотирозин, необходимый для этой цели. Единственным шансом сохранения структурной целостности молекулы тирозина в окружении аниона пероксинитрита является повышение концентрации SH-содержащих соединений, которые способны образовывать, взаимодействуя с анионом пероксинитрита, безопасные S-нитрозотиолы. Учитывая время полужизни аниона пероксинитрита, составляющее в фосфатном буфере при рН 7,4 и температуре 37 °С приблизительно 1 с, мы можем предположить, что он успевает диффундировать от места его образования на расстояние, равное нескольким клеточным диаметрам, что достаточно для того, чтобы активно участвовать в окислении липидов мембран, белков и химической модификации ДНК. Пероксинитрит окисляет остатки гуанина в ДНК с образованием нитрогуанина, который, в свою очередь, подвергается быстрой депуринизации с образованием в ДНК потенциально мутагенных апуриновых сайтов. Он вызывает также повреждение химической структуры ДНК путем образования 8-гидроксидезоксигуанозина. Таким образом, суммируя вышесказанное, мы можем заключить, что основные механизмы повреждающего действия аниона пероксинитрита заключаются в: 1) нарушении фосфорилирования тирозина; 2) нарушении функции белковых молекул; 3) усилении протеолиза белков; 4) инициации аутоиммунных реакций к гаптенам, возникающим в ходе нитрования белков. Пероксинитрит-индуцированное нитрование белков изменяет их конформацию вследствие снижения гидрофобности тирозина и/или путем изменения знака их заряда от нейтрального до отрицательного. Нитраты. С пищей и водой к нам в организм попадают нитраты. Нитраты для здоровья людей не опасны. Они малотоксичны и легко выделяются вместе с мочой, если человек пьет достаточное количество воды. У человека отсутствуют ферментные системы, метаболизирующие эти соединения, но бактерии, населяющие желудочно-кишечный тракт, способны восстанавливать нитраты в нитриты, которые являются токсичными для человека. Нитриты. В кислых условиях имеет место протонирование нитрита. В свою очередь азотистая кислота легко распадается с образованием окиси, диокиси и триокиси азота: NO3− → NO2− + H+ → HNO2 → NO• + NO2 + N2O3 . (39) Образовавшийся при диспропорционировании азотистой кислоты триоксид азота N2O3 может выступать донором иона нитрозония NO+ в реакциях нитрозилирования вторичных и третичных аминов с образованием канцерогенных N-нитрозаминов. В кислых условиях происходит протонирование азотистой кислоты до Н2NO2, образующей с амидами N-нитрозамиды. При нейтральных рН нитриты восстанавливаются до NO•. Таким образом, NO2−, легко восстанавливаясь в NO в организме млекопитающих, образует замкнутый метаболический цикл. Этот механизм начинает действовать особенно активно в условиях гипоксии. В свете представленных данных о механизмах токсического действия NO•/ONOO− становятся понятными токсические эффекты нитритов и нитратов. Токсическое действие нитритов в организме животных проявляется также в форме метгемоглобинемии. Нитрозилионы окисляют двухвалентное железо гемоглобина в трехвалентное. В результате такого окисления гемоглобин превращается в метгемоглобин, блокирующий центры переноса кислорода. В крови возрастает содержание молочной кислоты, холестерина, лейкоцитов, снижается количество белков. Особенно это состояние опасно для детей грудного возраста, у которых еще слабо работают ферментные системы. 6.1.4. Фосфор. К первоэлементам относится также фосфор. Его соединения важны для функционирования клетки. Вспомним, что фосфор входит в состав важнейших соединений: нуклеиновых кислот и нуклеотидов, фосфолипидов, коферментов и ферментов. Фосфор присутствует в клетке также в виде ионов ортофосфата и пирофосфата. Реакции фосфорилирования-дефосфорилирования определенных аминокислот в белках лежат в основе межклеточной химической сигнализации. Кости человека состоят из гидроксилапатита 3Са3(РО4)3·CaF2. В состав зубной эмали входит фторапатит. Фосфор вместе с магнием необходим для усвоения кальция, из которого построены костная и хрящевая ткани. Считают, что оптимальное соотношение содержания кальция и фосфора, так же как и кальция и магния, 2:1. Основную роль в превращениях соединений фосфора в организме человека и животных играет печень. Обмен фосфорных соединений регулируется гормонами и витамином D. Суточная потребность человека в фосфоре 800−1 500 мг. При недостатке фосфора в организме развиваются различные заболевания костей. Некоторые соединения фосфора: фосфин, зарин, диизопропилфторфосфат − очень токсичны. Зарин − изопропиловый эфир метилфторфосфоновой кислоты −
является боевым отравляющим веществом нервнопаралитического действия, вызывает поражение при любом виде воздействия, особенно быстро − при ингаляции. Зарин необратимо ингибирует фермент ацетилхолинэстеразу путём формирования ковалентного аддукта с аминокислотным остатком серина в активном центре фермента, что делает невозможным гидролиз ацетилхолина, медиато- ра, благодаря выбросу которого в межсинаптическое пространство синапса производится передача импульса к мышце или органу. В результате содержание ацетилхолина в межсинаптическом пространстве постоянно растёт, импульсы непрерывно передаются, поддерживая все иннервируемые вегетативными и двигательными нервами органы в активном состоянии вплоть до их полного истощения. Еще одно фосфорорганическое соединение − диизопропилфторфосфат − также ингибирует ферменты, содержащие остаток серина в активном центре, в том числе и ацетилхолинэстеразу, катализирующую следующую реакцию:
Реакция происходит каждый раз после проведения нервного импульса, прежде чем второй импульс будет передан через синапс. Диизопропилфторфосфат, одно из отравляющих веществ нервнопаралитического действия, как и зарин, образуя ковалентный аддукт с остатком серина в активном центре фермента, приводит к утрате способности нейронов проводить нервные импульсы, так как модифицированный остаток серина не может более ацетилироваться ацетилхолином (рис. 6.8).
6.1.5. Сера. В виде органических и неорганических соединений сера постоянно присутствует во всех живых организмах и является важным биогенным элементом. Биологическая роль серы определяется тем, что она входит в состав широко распространенных в живой природе соединений: аминокислот (метионин, цистеин, цистин, гомоцистеин), белков и пептидов; коферментов (кофермент А, липоевая кислота, S-аденозилметионин (SAM)), витаминов (биотин, тиамин), глутатиона и др. Сульфгидрилъные группы (-SH) остатков цистеина играют важную роль в структуре и каталитической активности многих ферментов. Образуя дисульфидные связи (-S-S-) внутри отдельных полипептидных цепей и между ними, эти группы участвуют в поддержании пространственной структуры молекул белков. У животных сера обнаружена также в виде органических сульфатов и сульфокислот − хондроитинсерной кислоты (в хрящах и костях), таурохолиевой кислоты (в желчи), гепарина, таурина. В некоторых железосодержащих белках (например, ферродоксинах) сера обнаружена в форме кислотолабильного сульфида. Сера способна к образованию богатых энергией связей в макроэргических соединениях, поэтому в реакциях синтеза в клетке многие соединения подвергаются активации путем образования тиоэфиров с коферментом А. Окислительно-восстановительный потенциал клетки во многом определяется соотношением тиол/дисульфид (вспомним многочисленные реакции восстановления, в которых принимает участие глутатион). Неорганические соединения серы в организмах высших животных обнаружены в небольших количествах, главным образом в виде сульфатов (в крови, моче), а также роданидов (в слюне, желудочном соке, молоке, моче). Морские организмы богаче неорганическими соединениями серы, чем пресноводные и наземные. Для растений и многих микроорганизмов сульфат (SO42-) наряду с фосфатом и нитратом служит важнейшим источником минерального питания. Перед включением в органические соединения сера претерпевает изменения в валентности и превращается затем в органических форму в своем наименее окисленном состоянии; таким образом сера широко участвует в окислительно-восстановительных реакциях в клетках. В клетках сульфаты, взаимодействуя с АТР, превращаются в активную форму − 3’-фосфоаденозил-5’-фосфосульфат (РАРS).
Катализирующий эту реакцию фермент − АТР:сульфат-адени- лилтрансфераза − широко распространен в природе. В такой активированной форме сульфонильная группа подвергается дальнейшим превращениям − переносится на другой акцептор или восстанавливается. Животные усваивают серу в составе органических соединений. Автотрофные организмы получают всю серу, содержащуюся в клетках, из неорганических соединений, главным образом в виде сульфатов. Способностью к автотрофному усвоению серы обладают высшие растения, многие водоросли, грибы и бактерии. Из культуры бактерий был выделен специальный белок, осуществляющий перенос сульфата через клеточную мембрану из среды в клетку. Большую роль в круговороте серы в природе играют микроорганизмы − десульфурирующие бактерии и серобактерии. Сера входит в состав антибиотиков (пенициллины, цефалоспорины); ее соединения используются в качестве радиозащитных средств, средств защиты растений. 6.1.6. Щелочные металлы: натрий и калий. Щелочные металлы в виде различных соединений входят в состав тканей живых организмов, преимущественно они находятся в виде иона Ме1+. Натрий и калий – жизненно необходимые макроэлементы, участвующие в обмене веществ организмов. Литий, цезий и рубидий, функции которых в организме мало изучены, можно отнести к примесным ультрамикроэлементам. Натрий содержится во внеклеточных жидкостях, а калий сосредоточен главным образом в клетках. Натрий и калий – антагонисты. При увеличении количества натрия в организме усиливается выведение калия почками, т. е. наступает гипокалиемия. Местом депонирования иона натрия в организме является костная ткань. Около 40 % натрия, содержащегося в костной ткани, участвует в обменных процессах, и благодаря этому скелет является либо донором, либо акцептором ионов натрия, что способствует поддержанию постоянства концентрации ионов натрия во внеклеточной жидкости. Ионы натрия играют важную роль: 1) в поддержании осмотического давления; 2) в обеспечении кислотно-основного равновесия организма; 3) в регулировании водного обмена; 4) в работе ферментов; 5) в передаче нервных импульсов; 6) в работе мышечных клеток. Хлорид натрия служит источником для образования Значение ионов калия важно в следующих физиологических процессах: 1) при сокращении мышц; 2) в нормальном функционировании сердца; 3) при проведении нервных импульсов; 4) в обменных реакциях; 5) в активации ферментов. В последние годы было показано, что моновалентные ионы выступают активаторами большого количества ферментов, при этом ион Na+ активирует ферменты, локализованные во внеклеточном пространстве, а ион К+ действует на внутриклеточные белки. В ферментативных системах эти ионы координируются с 5−8 лигандами, чаще всего встречается октаэдрическая координация:
Действие моновалентных катионов на ферменты может реализовываться двумя способами: кофакторно или аллостерически. В первом случае, когда катион выступает кофактором в ферментативной реакции, реакция не протекает в его отсутствие. Как правило, в этом случае моновалентный катион осуществляет свое активирующее воздействие на фермент совместно с дивалентным катионом, например Mg2+ или Mn2+. Во втором способе активатор непосредственно не связывается с субстратами, но оказывает влияние на ферментативную реакцию путем изменения конформации белка после связывания с аллостерическим центром.
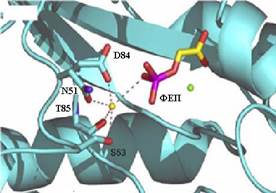
Рис. 6.9. Пространственная структура активного центра пируваткиназы из дрожжей. Ион К+ изображен желтым шариком, ион Mn2+ − зеленым, фосфатная группа фосфоенолпирувата (ФЕП) − красно-лиловым. Ионы К+ и Mn2+ оптимизируют связывание субстрата и облегчают перенос фосфатной группы от ФЕП на ADP (согласно [25]) Так, пируваткиназа и фосфофруктокиназа относятся к К+-активи-руемым ферментам I типа. На рис. 6.9 представлен активный центр пируваткиназы, где видно, что совместное воздействие ионов К+ и Mn2+ облегчает перенос фосфатной группы от фосфоенолпирувата к ADP. Некоторые факторы свертывания крови (фактор Xа, тромбин) являются Na+-активируемыми ферментами II типа. Например, в тромбине ион Na+ связывается в аллостерическом сайте вблизи от активного центра, изменяя конформацию расположенного рядом участка белка таким образом, что каталитический Asp189 ориентируется относительно Arg, пептидная связь которого должна гидролизоваться (рис. 6.10).
Поиск по сайту: |
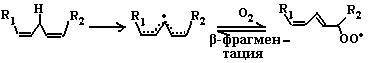
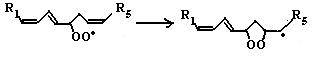 (7)
(7)



 (21)
(21) (22)
(22)


 (23)
(23) (26)
(26) (27)
(27)
 (28)
(28)
 (31)
(31)
 (32)
(32)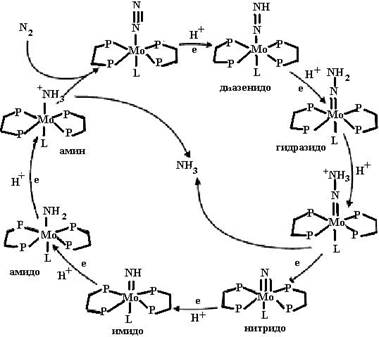
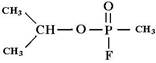

 Рис. 6.8. Действие диизопропилфторфосфата на ацетилхолинэстеразу (АХЭ)
Рис. 6.8. Действие диизопропилфторфосфата на ацетилхолинэстеразу (АХЭ)
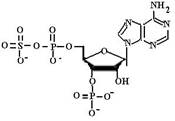
 (40)
(40)