|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Саморегуляция поведения
Пока еще на этот вопрос нельзя дать исчерпывающего ответа. Но некоторые указания относительно такого механизма можно получить, анализируя взаимоотношения между лобной корой и лимбической системой, а также процесс решения задач с отсроченным чередованием и с отсроченной реакцией. Ключ к правильному выполнению теста с чередованием (который сопровождается коррекцией, то есть повторяется до тех пор, пока не будет дан правильный отчет) можно найти в недавно завершенном опыте. Обезьяна должна была составить простой стратегический план, который учитывает контекст, созданный непосредственно предшествовавшим опытом. В этом случае ей очень помогает стратегия «перемещение — ответ». Однако следует отметить, что правильность поведения в любой момент зависит не от событий, происходящих в этот момент в окружающей среде, а от контекста, — созданного памятью (кратковременной). В главах XV мы выяснили, что амигдала и гиппокамп, входящие в лимбическую систему, регистрируют и оценивают поведение. Поэтому не удивительно, что при повреждении этих структур организм неспособен выполнить задачу, в которой правильность ответа зависит от того, зарегистрирована ли (и оценена ли) предыдущая задача. Чтобы создать стратегический план, связанный с контекстом, контекст должен быть сформирован и сохранен. Однако дефекты выполнения задач на чередование после лобной резекции можно объяснить по-другому. Контекст устанавливается, но организм, очевидно, не знает, как им пользоваться. Мы столкнулись с этой проблемой при объяснении результатов удаления задней «ассоциативной» коры. В этом случае (Bagshaw, Mackworth, Pribram, 1971) запись движений глаз показала, что — обезьяны нормально рассматривали объект и, очевидно, хорошо видели сенсорные сигналы, но не могли действовать на основе того, что они воспринимали. У больных отчетливо проявляется эта неспособность регулировать поведение, когда смысл его зависит от контекста, в котором протекает их деятельность. Клинические наблюдения изобилуют примерами неадекватного поведения. Возможно, наиболее показательным из них является наблюдение, проведенное над одним из больных Г. Л. Тёйбера (1964). Этот больной регулярно посещал лабораторию по четвергам. Когда один из четвергов совпал с праздником, он, как обычно, отправился в лабораторию, говоря себе на каждом шагу: «Сегодня праздник, мне не нужно идти в лабораторию, там никого нет». Специальный контекст «праздника», хотя и вербализованный, не оказывал влияния на его поведение. Только когда он дошел до лаборатории и не обнаружил там никого, он вернулся домой, однако так и не сделав вывода о происшедшем.
Мы уже много говорили по поводу задачи на чередование. Нарушение, проявляющееся в классической прямой и косвенной формах задач на отсроченную реакцию после резекции лобных долей мозга, объясняет нам еще кое-что относительно взаимоотношений между восприимчивостью к контексту и кратковременной памятью. В этом случае правильность выполнения задания зависит не от контекста, установленного в результате какого-то предшествовавшего действия, а от временного появления сигнала, указывающего на начало пробы. Множество экспериментов показало, что правильная регистрация мозгом этого на мгновение появляющегося знака, способствующая его усвоению путем активной реакции на сигнал, является решающим условием верного выполнения задания (Pribram, 1969b). Что же включает в себя правильная регистрация? В нейрофизиологическом отношении, если нет некоторых изменений в электрической активности мозга, это приведет к ошибке. Именно такие изменения указывают на появление ориентировочной реакции и образуют основу для последующего привыкания. И действительно ориентировочная реакция резко изменяется под влиянием поражений лобных долей как у человека, так и обезьяны (Лурия, Pribram, Хомская, 1964). Теперь уже хорошо известно, что имеется ряд индикаторов ориентировочной реакции (Е. Н. Соколов). К ним относится поворот глаз к стимулу, изменение частоты сердцебиений и дыхания, КГР, изменение кровенаполнения — периферических и центральных сосудов (измеряемое при помощи пальцевой и височной плетизмографии) и активация в ЭЭГ. После резекции лобных долей мозга (см. Рис. XVIII—7), так же как после амигдалэктомии, остаются только поведенческие и электроэнцефалографические индикаторы (хотя продолжительность активации в ЭЭГ сокращается).
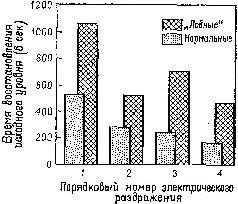
Вместе с тем повторение сигнала в этих случаях уже не вызывает привыкания и поведенческие реакции продолжают сохраняться. Как уже подробно говорилось, это привело к предположению, что ориентировочная реакция состоит по крайней мере из двух отдельных процессов: из компонента настораживания, поиска и отбора стимулов и из их регистрации в сознании и памяти. Лобная кора и амигдала оказывают влияние на этот процесс регистрации. Когда регистрация нарушается, организм не только утрачивает способность припоминать события, но и становится в гораздо большей степени подверженным ретроактивному и проактивному торможению, обусловленному интерферирующими воздействиями. Люди и обезьяны с лобными повреждениями чувствительны к интерференции, влияющей на процессы кратковременной памяти (Crueninger and Pribram, 1969; Pис. XVIII—8). Данные электрофизиологии говорят о том, что лобная кора в норме контролирует интерференцию посредством кортикофугальных влияний на афферентные системы. Электростимуляция лобной коры вызывает результаты, противоположные эффектам стимуляции задней ассоциативной коры (см. Рис. XVIII—9). Раздражение лобных отделов мозга увеличивает избыточность каналов (см. Рис. XI—6 и Х1—7; каналы имеют тенденцию в любой момент действовать все вместе. Таким образом, возможность интерференции между следами последовательных сигналов сокращается. Б проводившихся экспериментах длительность вызванного изменения равнялась миллисекундам; ретро- и проактивная интерференция в поведенческих ситуациях занимает значительно большее время. Тем не менее приведенные электрофизиологические данные наводят на мысль, что, по-видимому, в поведенческих ситуациях, в которых наблюдаются эффекты интерференции, лобная кора обеспечивает более длительное сохранение возбуждения, чем нам удавалось достигнуть в эксперименте. Приведенный анализ опытов с отсроченным чередованием и отсроченными реакциями позволяет предположить, что повреждение лобных долей вызывает нарушение тех процессов мозга, существенным элементом которых является кодирование изменений состояний. Такие процессы свойственны работе кратковременной памяти, предполагающей скорее решения, зависящие от контекста, чем хорошо упроченные действия, не зависящие от контекста, и нарушения их отражаются как на решении задач, так и на эмоциональном поведении. Можно предположить, что подобный дефект сказывается на решении задач потому, что организм не в состоянии регулировать свое поведение на основе событий, вносящих беспокойство и сигнализирующих об изменениях в контексте. Этот дефект проявляется в эмоциональном поведении: организм не справился с контролем, регистрацией и оценкой изменений, непрерывно усложняющих контекст и таким образом увеличивающих беспокойство в настоящем. При повреждении височной доли утрата способности к восприятию контекста проявляется в том, что поток событий не сохраняется в следах и, таким образом, протекает только в настоящем, которое не имеет ни прошлого, ни будущего. Организм становится автоматом, находящимся во власти своих мгновенных состояний, которые он не может регулировать.
Поиск по сайту: |