|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Гладкая эндоплазматическая сетьСтр 1 из 2Следующая ⇒
Лекция 3. Вакуолярная система План лекции
Определение вакуолярной системы Вакуолярная система - это система органоидов, состоящих из мембранных пузырей разной формы, определенным образом связанных друг с другом и плазматической мембраной.
Одно из существенных свойств вакуолярной системы - разделение клетки на отсеки (компартменты) - гиалоплазму и содержимое внутри мембранных отсеков.
В состав вакулярной системы входят следующие компоненты: шЭПС, глЭПС, кГ, лизосомы и СЯО.
Эндоплазматическая сеть (ЭПР) Эндоплазматическая сеть состоит из двух разновидностей - гладкой и шероховатой, которые отличаются отсутствием или наличием на поверхности мембран рибосом. Этот органоид относится к органоидам общего назначения и входит в состав цитоплазмы всех типов клеток эукариот. Шероховатая ЭПС Этот органоид был открыт в 1943 г. Claude методом дифференциального центрифугирования. При разделении клеточного гомогената на фракции в центрифужных пробирках можно идентифицировать 3 основные фракции: надосадочную жидкость, микросомную и ядерную фракции. Именно в состав микросомной фракции, которая содержит множество вакуолей с разнообразным содержимым, входят компоненты вакуолярной системы.
Схема строения ЭПС гепатоцита (рис. Пунина М.Ю.) 1 – шероховатая ЭПС; 2 – гладкая ЭПС; 3 - митохондрия В 1945 г. Porter при изучении в электронном микроскопе целых клеток куриных фибробластов обнаружил у них в зоне эндоплазмы мелкие и крупные вакуоли и соединяющие их канальцы. Именно этот компонент клетки и был назван эндоплазматической сетью. При помощи методов просвечивающей электронной микроскопии было установлено, что ЭПС состоит: ·
Рис. Эндоплазматическая сеть 1 – трубочки гладкой ЭПС; 2 – цистерны гранулярной (шероховатой) ЭПС; 3 – наружная ядерная мембрана, покрытая рибосомами; 4 – поровый комплекс; 5 – внутренняя ядерная мембрана (по Кристичу с изменениями).
Эти мембранные мешки, как видно на электронномикроскопических фотографиях концентрическими слоями сосредоточены вокруг ядра. Размер внутреннего отсека составляет около от 20 нм до 1 мк (1 000 нм). Количество элементов шЭПС зависит в клетках от их функции и степени дифференциации. Сосредоточение цистерн шЭПС в клетках в области вокруг ядра называется эргастоплазмой и свидетельствует об участии таких клеток в синтезе экспортного белка. Рибосомы, прикрепленные к поверхности мембран шЭПС, могут быть единичными, так и в виде розеток (полисом). Глубина проникновения рибосом внутрь мембран также может отличаться.
Механизм функционирования шероховатой .ЭПС 1. Функция синтеза экспортного белка. Гипотеза Блобеля и Сабатини (1966 - 1970). Эта функция осуществляется при участии самих мембран шЭПС и примембранного слоя гиалоплазмы, в котором сосредоточена система, отвечающая за все этапы трансляции. Предполагается, что на поверхности мембран шЭПС имеются специальные участки, отвечающие за узнавание концевых фрагментов молекул иРНК. Прикрепление этих молекул предшествует началу собственно процесса трансляции. В ходе трансляции, синтезируемые экспортные белки проникают сначала через канал в большой субъединице рибосомы, а затем и через мембрану. Внутри мембранного отсека эти белки накапливаются. Их дальнейшая судьба связана с процессами дозревания. 2. Сегрегация и преобразование экспортных белков. Сущность процессов дозревания заключается в том, что у отдельных белковых молекул при помощи специальных ферментов отрезается сигнальная последовательность, другие ферменты присоединяют к ним либо радикалы, либо фрагменты углеводных и липидных молекул, в случае формирования сложных по химическому составу секретов. В случае, если это белки мембран, то в зависимости от их положения в билипидном слое (снаружи, внутри или на поверхности молекулы белков перемещаются из большой субъединицы рибосомы на ту или иную поверхность мембраны или пронизывают ее насквозь (интегральные белки).
Схема молекулярной оргаизации шероховатой ЭПС и ее роли в процессах синтеза и вторичных преобразований белковых молекул (рис. Пунина М.Ю.) 1 – мембрана; 2 – полуинтегральные белки и гликопротеиды; 3 – олигосахариды и другие углеводные компоненты на внутренней поверхности мембран и в полости цистерн; 4 – иРНК; 5 – гипотетический рецептор в мембране для иРНК; 6, 7 – субъединицы рибосом; (6 – малая, 7 – большая); 8 – неиндетифицированные интегральные белки мембраны, обеспечивающие прохождение синтезируемых белков через мембрану; 9 – гипотетические интегральные белки, обеспечивающие крепление к мембране больших субъединиц рибосом; 10 – синтезируемая белковая молекула; 11 – 13 – варианты синтеза интегральных (13), полуинтегральных белков наружного (11), и внутреннего (12) слоев мембраны; 14 – синтез белков гиалоплазмы на прикрепленной рибосоме; 15 – 17 – последовательные стадии синтеза, прохождения через мембрану и вторичных изменений экспортных белков. В левом верхнем углу - внешний вид шероховатой ЭПС в электронном микроскопе; в правом углу – типичные отношения между полисомой и мембраной шероховатой ЭПС при синтезе экспортных и полуинтегральных белков; в центре – цитоплазматический пул субъединиц рибосом . Стрелки показывают направление перемещения субъединиц рибосом и синтезированных белковых молекул. 3. Внутримембранное хранение веществ. Некоторые секреты хранятся во внутри мембранном пространстве определенное время, по прошествии которого они упаковываются в мелкие мембранные пузырьки, которые переносят секрет от шЭПС в зону формирования комплекса Гольджи. Так при изучении образования белковых молекул антител было установлено, что сама молекула строится за 90 сек, но снаружи клетки она оказывается только через 45 минут. То есть при секреции установлены следующие этапы: синтез белка, сегрегация (разъединение), внутри клеточный транспорт, концентрирование, внутриклеточное хранение, освобождение из клетки. 4. Участие в обновлении мембранных компонентов (место образования новой мембраны). Гипотеза Лодиша и Ротмена (1977). Внутренняя часть билипидного слоя мембранных цистерн шЭПС – место встраивания вновь синтезированных молекул липидов. После нарастания поверхности внутренней части билипидного слоя избыток липидных молекул перескакивает в наружный слой билипидной поверхности из-за подвижности липидных молекул по вертикали (свойство флип-флоп).
Гладкая эндоплазматическая сеть В отличие от шЭПС эта разновидность сети имеет два существенных отличия: · мембранные пузыри имеют форму сложной системы трубочек; · поверхность мембраны гладкая, лишена рибосом.
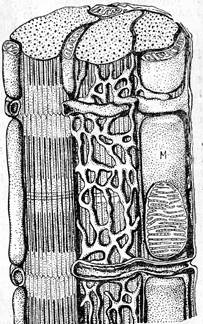
Схема расположения трубочек гладкой ЭПС (саркоплазматического ретикулюма) мышц. М – митохондрии. (по Fawcett, McNutt, 1969) Этот органоид также относится к органоидам общего назначения, но в некоторых клетках составляет основную массу цитоплазмы таких клеток. Это связано с тем, что эти клетки участвуют в образовании не мембранных липидов. Примером таких клеток служат клетки коры надпочечников, специализирующихся на выработке стероидных гормонов. В цитоплазме этих клеток наблюдается сплошная масса трубочек гладкой ЭПС. Гладкая ЭПС обычно занимает в клетке строго определенное место: в клетках кишечника – в апикальной зоне, в клетках печени в зоне отложения гликогена, в интерстециальных клетках семенника она равномерно распределена по всему объему цитоплазмы. Происхождение гладкой ЭПС – вторичное. Этот органоид образуется из шЭПС в результате утери последним рибосом, либо за счет роста шЭПС в виде трубочек, лишенных рибосом..
Механизм функционирования гладкой ЭПС 1. Участие в синтезе не мембранных липидов. Эта функция связана с секрецией этих веществ, например стероидных гормонов. 2. Детоксикация (внутри мембранное хранение токсических отходов метаболизма). Эта функция связана со способностью трубочек гладкой ЭПС клеток печени накапливать во внутри мембранном пространстве ядовитых продуктов метаболизма, например некоторых лекарств (явление известное для барбитуратов). 3. Накопление двухвалентных катионов. Эта функция характерна для L-каналов мышечных волокон. Внутри этих каналов накапливаются двухвалентные ионы Ca+2,которые участвую в процессах образования кальциевых мостиков между молекулами актина и миозина в процессе мышечного сокращения.
Поиск по сайту: |
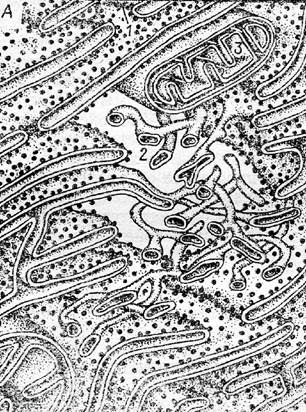
 из системы плоских мембранных мешков (цистерн), соединенных перемычками (анастомозами).
из системы плоских мембранных мешков (цистерн), соединенных перемычками (анастомозами).