|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Водно-солевой обмен на суше
Приспособления к водообеспечению у растений влажных местообитаний Растения, обитающие во влажных биотопах, подразделяются на несколько экологических групп: 1. Гидрофиты – наземные растения, укореняющиеся в дне водоема или в переувлажненной почве. К ним относятся: тростник, кубышка, калужница, вахта и др. Приспособления к условиям среды у гидрофитов следующие: 1) хорошо выраженная аэренхима и общая губчатость тканей – является приспособлением к недостатку кислорода в переувлажненной почве; 2) погруженные в воду части растений имеют полости, сообщающиеся с устьицами находящихся в воздухе частей, что способствует газообмену; 3) отсутствие кутикулы и перидермы у подводных органов – способствует получению воды и минеральных солей, минуя корневую систему. 2. Гидафиты – растения, тело которых полностью погружено в воду. К ним относятся: элодея, ряска, рдесты и др. Приспособлением к данным условиям среды является редукция корневой системы. Поглощение воды и минеральных солей происходит через всю поверхность тела. 3. Гигрофиты – наземные растения, обитающие в местах с высокой влажностью воздуха и хорошим почвенным водоснабжением. К ним относятся растения влажных тропических лесов и тенистых лесов умеренной зоны. Гигрофиты делятся на теневые (произрастают во влажном ярусе тенистых лесов) и световые (произрастают на открытых местах в условиях высокой влажности). Приспособлением к условиям среды у теневых гигрофитов является наличие особых водяных устьиц, выделяющих капельно-жидкую воду. Приспособлением у световых гигрофитов является более эффективная транспирация. 3. Мезофиты – широкий круг растений, занимающих различные местообитания, в том числе не сильно увлажненные. Мезофиты подразделяют на пять групп: 1) вечнозеленые – деревья и кустарники влажных тропиков; 2) зимне-зеленые деревянистые – растения тропиков и субтропиков, сбрасывающие листву и впадающие в неактивное состояние в сухие периоды; 3) летне-зеленые деревянистые – деревья и кустарники умеренной зоны, сбрасывающие листву и впадающие в неактивное состояние в зимнее время; 4) летне-зеленые травянистые – растения, у которых наземные части, кроме почек возобновления, отмирают к зиме; 5) эфемеры и эфемероиды – растения, обитающие в аридных (засушливых) зонах, которые вегетируют в течение короткого влажного периода. Приспособления у мезофитов к ограниченному водоснабжению и изменчивой влажности воздуха следующие: 1) Высокое осмотическое давление клеточного сока, обеспечивающее сосущую силу корневой системы. 2) Впадение в неактивное состояние в неблагоприятные периоды. 3) Структура корневой системы двух типов: экстенсивная и интенсивная. Экстенсивная охватывает большой объем почвы, но слабо ветвится в нем. Интенсивная охватывает небольшой объем почвы, но сильно ветвится и имеет множество корневых волосков. Водный обмен и осморегуляция у земноводных
Большинство земноводных (амфибий) пресноводны. Поэтому водно-солевой обмен у них регулируется способом, сходным с таковым у пресноводных рыб. Структура и функции почек у амфибий такая же, как у пресноводных рыб: она обеспечивает большие потери воды с мочой. Примитивные легкие земноводных не обеспечивают достаточный уровень газообмена, поэтому в дыхании участвует кожа. Это определяет структуру кожи – она голая и слизистая. Кожа амфибий легко проницаема и для воды. Через такую кожу происходят большие потери воды, поэтому большинство земноводных ведет околоводный образ жизни. При нахождении амфибий в воде и при достаточном увлажнении вода легко проникает через кожу – так компенсируются потери воды, а избыток воды выводится в составе мочи. Ионы Na+ и Сl – реабсорбируются в почках, а, кроме того, путем активного переноса проникают через кожу. Приспособления к дефициту влаги у амфибий следующие: 1. Запасание воды в составе мочи в мочевом пузыре. При необходимости эта вода может возвращаться в кровеносное русло. 2. Морфологические приспособления, ограничивающие потери воды через кожу. К ним относится слизь, покрывающая кожу, которую продуцируют железистые клетки (рис.3). Слизь замедляет испарение воды. К морфологическим приспособлениям, ограничивающим потери воды, относятся также утолщения кожного покрова – «бородавки». 3. Морфологические приспособления, участвующие в восполнении водного дефицита – специализированные участки кожи, обеспечивающие проникновение влаги с поверхности почвы и росы с растений. Например, участки кожи с более тонким однослойным эпителием. 4. Адаптивное поведение. Многие амфибии приурочивают время активной деятельности к периодам наибольшей влажности (например, наибольшая активность ночью или после выпадения дождей).
Рис. 3. Схема разреза через кожу земноводных: 1 – эпидермис, 2 – кориум, 3 – пигментные клетки, 4 – железистые клетки, 5 – мускульная оболочка кожной железы, 6 – выводной проток железы. Водно-солевой обмен в засушливых местообитаниях Приспособления растений к засушливым местообитаниям Растения, приспособившиеся к обитанию в местах с недостаточным увлажнением, относятся к группе ксерофитов. По принципу адаптаций к данным условиям среды они делятся на две группы: 1. Суккуленты. Главные принципы адаптаций у суккулентов – запасание воды и экономное использование воды. К суккулентам относятся кактусы, агавы, алоэ и др. Приспособления к запасанию и экономному использованию воды следующие: 1) Хорошо развитая водозапасающая паренхима в листьях и стеблях. Суккуленты имеют сочные и мясистые органы. 2) Интенсивное насасывание влаги осадков из самых верхних горизонтов почвы (из-за низкого осмотического давления клеточного сока использование глубокой почвенной влаги для них недоступно). 3) Высокое содержание сахаров в клеточном соке, повышающих его водоудерживающую силу. 4) Экономное испарение. Экономному выведению воды испарением способствует редукция листьев, превращение их в колючки, кутикула на поверхности листьев, восковой налет или густое опушение. Кроме того, суккуленты имеют погруженные устьица, которые днем закрыты, что снижает потери воды. Устьица открываются ночью, в это время поглощается СО2, который используется в фотосинтезе на следующий день. 3. Склерофиты – растения, которые не способны запасать воду в органах и тканях. Они выглядят как сухие и жесткие растения. Принципом адаптации к засушливым условиям у склерофитов является способность к активной перестройке водного обмена в зависимости от обеспеченности влагой. Приспособления к аридности климата у склерофитов следующие: 1) Высокая тканевая устойчивость к дегидратации. Эти растения могут терять до 25 % влаги без патологических последствий, так как их цитоплазма сохраняет свои свойства даже при такой степени обезвоживания, которая смертельна для других организмов. 2) Большая сосущая сила корней. Это определяется высоким осмотическим давлением клеточного сока и позволяет извлекать влагу даже при малом содержании ее в почве. Приспособления растений к засолению почвы
Почвы аридных зон часто содержать повышенное количество солей. Солеустойчивость разных видов растений различна. Для многих растений избыточное количество солей вредно, так как нарушает клеточно-тканевые процессы. Однако существуют растения, которые переносят высокие концентрации солей и могут произрастать на засоленных почвах. Это галофиты. Галофиты не просто устойчивы к засоленности почвы, но и испытывают необходимость в высокой солевой нагрузке. Приспособления к условиям засоленных почв у галофитов следующие: 1. Высокое осмотическое давление клеточного сока (благодаря высокой концентрации солей, а также некоторых других веществ, например, углеводов). Это ведет к увеличению сосущей силы корней, более эффективному насасыванию воды из почвы. 2. Слабая проницаемость клетки для электролитов. В результате избыток солей не наносит вреда цитоплазме клеток. Однако на засоленных почвах произрастают не только галофиты, но и растения, для которых избыток солей веден. Принципом адаптации таких растений к засоленности является выведение избытка солей. Некоторыми примерами приспособлений таких растений являются следующие: 1) выведение избытка солей через специальные железы на поверхности листа; 2) связывание избытка солей с органическими веществами протопласта; 3) концентрация солей в отдельных участках органов (например, в особых волосках на листьях) и выведение их, таким образом, из метаболизма. Водный обмен у наземных животных. Приспособления к аридности климата Наземная (воздушная) среда обитания в аридных зонах отличается низкой или колеблющейся влажностью воздуха. Это создает постоянную угрозу обезвоживания для наземных животных. Основными принципами адаптации к данным условиям для животных являются: 1) экономия водных ресурсов; 2) эффективное использование метаболической воды; 3) повышение надежности способов добычи воды из внешней седы. В наземной среде у животных происходит разделение водного и солевого обмена (связь остается только на уровне метаболических процессов). Имеются определенные особенности поступления и выведения воды и минеральных веществ: 1. Минеральные вещества не поступают через кожные покровы, как в водной среде. Они попадают в организм только с пищей и выводятся в составе мочи и фекалий. 2. Вода не поступает в организм и не удаляется путем осмотических механизмов, а попадает только вместе с пищей или путем питья. Эти особенности определили сходство водного и солевого обмена у разных групп наземных животных. У наземных беспозвоночных (насекомых и паукообразных) имеются следующие приспособления к аридности климата: 1. Слабая проницаемость покровов для воды, что снижает потери воды путем испарения. Наземные беспозвоночные имеют плотные хитиновые покровы, покрытые тончайшим восковым слоем, который препятствует испарению (рис.4). Сравните потери воды путем испарения у насекомых и других животных (табл.2). Таблица 2. Потери воды с поверхности тела при комнатной температуре
3. Преобразованная выделительная функция. Моча формируется в мальпигиевых трубочках секреторным путем. Моча и фекалии попадают из мальпигиевых трубочек в заднюю кишку, где вода интенсивно абсорбируется, и продукты выделения выводятся практически в обезвоженном состоянии. 4. Повышенная способность к утилизации метаболической воды. Это свойство у некоторых групп выражено настолько отчетливо, что многие из них могут существовать в условиях полного отсутствия воды, восполняя потребности в ней только метаболическим путем. У позвоночных наиболее совершенные адаптации к жизни в воздушной среде сформировались у пресмыкающихся, птиц и млекопитающих. Приспособления к аридности климата сходны с таковыми у наземных беспозвоночных, хотя и имеют свои особенности: 1. Слабая проницаемость покровов для воды. Это достигается тем, что наружные слои клеток эпидермиса заполнены роговым веществом, которое препятствует испарению воды с поверхности тела (табл.3). 2. Отсутствие кожных желез у пресмыкающихся и птиц, что также уменьшает кожное испарение (рис.6).
Рис. 4. Продольный разрез кожи ящерицы: 1 – эпидермис, 2 – собственно кожа (кориум), 3 – роговой слой, 4 – мальпигиев слой, 5 – пигментные клетки, 6 – кожные окостенения.
3. Повышенная способность к образованию и использованию метаболической воды. Наибольшее количество метаболической воды выделяется при окислении жиров (1,05-1,07 г на 1 г окисленного жира). Многие животные пустынь перед наступлением засушливого сезона накапливают большие запасы жира и расходуют их во время летней спячки или оцепенения для обеспечения потребности в воде. Так как в таком состоянии общий уровень метаболизма резко снижен, этих запасов хватает на длительное время.
Таблица 3. Потери воды с поверхности тела при комнатной температуре у некоторых видов рептилий
4. Преобразованная выделительная функция. У наземных позвоночных имеются тазовые почки, в которых в отличие от почек рыб отсутствуют воронки, открывающиеся в полость тела. Такая структура почек обеспечивает резкое снижение водных потерь путем отключения пассивного выведения воды из полости тела. У млекопитающих, кроме того, в почках имеется петля Генле – мощный концентрационный аппарат, позволяющий извлекать из первичной мочи большую часть воды (рис.5). Поэтому в условиях дефицита влаги выводится моча высокой концентрации.
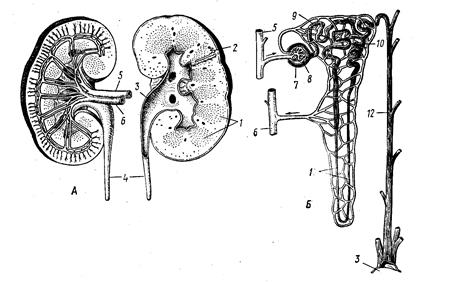
Рис. 5. Почка млекопитающих. А – разрез через почку человека; Б – схема циркуляции крови в почке:
1 – почечные пирамиды, 2 – положение нефрона, 3 – почечная лоханка, 4 – мочеточник, 5 – почечная артерия, 6 – почечная вена, 7 – боуменова капсула, 8 – сосудистый клубочек, 9 – проксимальный и 10 – дистальный извитые отделы почечного канальца, 11 – петля Генле, 12 – собирательный каналец. 5. Кишечная реабсорбция. В толстой кишке происходит обратное всасывание воды благодаря осмотическому току воды через слизистую оболочку. При дегидратации организма этот процесс ускоряется за счет увеличения осмотического градиента. 6. Адаптивное поведение – активный выбор местообитаний вблизи водоемов или с благоприятным микроклиматом (например, с высокой влажностью и относительно низкой температурой). Солевой обмен у наземных позвоночных Для обитателей аридных областей важна не только экономия воды, но и выведение избытка солей, который возникает при водном дефиците. Однако в природных условиях могут возникать и ситуации, характеризующиеся солевой недостаточностью. Поэтому у наземных позвоночных имеется два вида адаптаций: 1) направленные на выведение избытка солей; 2) направленные на восполнение минералов. Приспособления, направленные на избавление от избытка солей, включают в себя выведение солей через: 1. Почки. Степень развития этой функции зависит от экологической специализации вида. 2. Кишечник. Обезвоживание организма усиливает выведение зольных веществ с пометом. 3. Солевые железы. В условиях солевой нагрузки в них образуется концентрированный солевой раствор, который выводится наружу. Адаптации, направленные на восполнение недостатка солей, включают в себя адаптивное поведение, которое заключается в поедании пищи, содержащей минеральные элементы, а именно: скорлупы яиц, костей павших животных, гальки, почвы, слизывание кристаллов соли с засоленной почвы.
Поиск по сайту: |