Преемственность наследственных свойств определяется универсальными эволюционно закрепленными процессами деления клеток.
Митоз обеспечивает преемственность в ряду клеточных поколений, а мейоз - в ряду поколений организмов.
В основе митоза и мейоза лежит фундаментальная способность хромосом к автокаталитической редупликации, в результате которой все расположенные по длине хромосомы гены удваиваются, а затем распределяются по дочерняя клеткам.
Клеточный и митотический циклы
Клеточный цикл - это жизненный цикл клетки, это период существования клетки от момента ее образования путем деления материнской клетки до ее собственного деления или смерти.
Обязательными компонентами клеточного цикла являются:
митотический (пролиферативный);
период выполнения клеткой в многоклеточном организме специальных функций;
период покоя, где судьба клетки неопределена (клетка либо начинает подготовку к делению, либо пойдет по пути специализации.
Клеточный цикл представляет собой сложную систему реакций и процессов идущих в строгой последовательности. Однако лишь немногие клетки способны к делению в течение длительного периода. Таковыми являются клетки меристем растений и обновляющихся тканей животных. У одноклеточных организмов клеточный цикл совпадает с жизнью особи, ее онтогенезом. У непрерывно размножающихся тканевых клеток (эмбриональных клеток) клеточный цикл совпадает с митотическим циклом.
Продолжительность клеточного цикла и его периодов составляет в размножающихся клетках 10 - 50 часов и зависит от ряда причин: типа клеток, возраста, гормонального баланса организма, количества ДНК в ядре, температуры, времени суток и др.
При этом митоз занимает 5 - 10% времени клеточного цикла. Клеточный цикл может длиться недели и месяцы (например, у клеток печени). Для нервных клеток Клеточный цикл равен продолжительности жизни организма.
Митотический цикл включает
подготовку клетки к делению или интерфазу (репродуктивная фаза);
собственно деление или митоз (разделительная фаза).
Продолжительность митотического цикла для большинства клеток 10 - 50 часов. У разных клеток стадии митотического цикла варьируют, например, у млекопитающих для митоза необходимо 1-1,5 часа, для синтетического периода - 6-10 часов.
Длительность интерфазы различна у разных типов клеток: некоторые имеют короткую интерфазу, другие, наоборот, большую часть жизни находятся в репродуктивной фазе.
Интерфаза состоит из 3-х периодов:
постмитотический (предсинтетический период) - G1. В этом периоде происходит:
интенсивный метаболизм под контролем ферментов;
интенсивные биосинтезы;
образование митохондрий, у растений - хлоропластов;
синтез ЭПС, лизосом, аппарата Гольджи вакуолей, пузырьков;
образование при участии ядрышка рРНК, синтез мРНК, тРНК;
образуются рибосомы;
идет активный синтез структурных и функциональных белков;
происходит интенсивный рост клетки;
происходит образование веществ подавляющих или стимулирующих начало новой фазы.
синтетический период - S. Здесь не происходит удвоение числа хромосом, удваивается количество нитей ДНК за счет редупликации ДНК. Процесс репликации основан на ряде принципов:
комплементарности - цепи материнской ДНК расходятся, и каждая из них служит матрицей, на которой достраивается комплементарная цепь;
полуконсервативности - в дочерних ДНК сохраняется (консервируется) в неизменном виде лишь половина материнской ДНК;
антипараллельности - синтез комплементарных цепей идет в противоположных направлениях;
прерывистости - ДНК эукариот содержит слишком много пар нуклеотидов и сразу "раскрыть" и вытянуть в линию всю молекулу невозможно, поэтому удвоение начинается в нескольких пунктах одновременно, путем образования нескольких репликативных вилок. Набор хромосом остается диплоидный - 2п, число нитей ДНК становится - 4с; синтезируются белки-гистоны, с ними связана каждая нить ДНК.
постсинтетический (предмитотический период) - G2 . Это самый короткий период интерфазы. Для него характерно:
интенсивный биосинтез фибриллярных белков - тубулинов, лежащих в основе строения микротрубочек;
деление путем почкования митохондрий, пластид;
накопление энергетических запасов;
репликация центриолей;
начало формирования веретена деления
Биологическое значение митотического цикла:
обеспечивает приемственность наследственной информации в ряду клеточных поколений
служит универсальным механизмом воспроизведения клеточной организации эукариотических клеток.
Главные события митотического цикла:
редупликация ДНК в синтетический период интерфазы;
равномерное распределение наследственного материала между дочерними клетками за счет расхождения дочерних хроматид в анафазе митоза.
Митоз
Митоз (кариокинез) - это процесс деления ядра. Обычно под митозом цитологи обьединяют "кариокинез" + "цитокинез". Название "митоз" происходит от греческого слова "митос" - нить. Митоз это - непрямое деление клетки, основной способ деления соматических клеток эукариот. К митотическому делению способны как диплоидные, так и гаплоидные клетки, но дочерние клетки в любом случае содержат набор хромосом, идентичный материнской. Число дочерних клеток при митозе всегда равно двум.
В 1874г. И. Д. Чистяков описал ряд фаз митоза в спорах плаунов. В 1876 -79г.г. М. Э. Страсбургер детально исследовал митоз в клетках растений. В 1882г. В.Флемминг исследовал митоз в клетках на животных.
Средняя продолжительность митоза 1 -2 часа. По частоте митозов ткани подразделяются на 3 группы:
стабильные ткани. К ней относят нервную ткань. Клетки нервной ткани делятся только в эмбриональном периоде;
ткани растущие. Здесь большинство клеток находятся в интерфазе, а часть делится (например, почки, поперечно-полосатая мышечная ткань);
обновляющиеся ткани. Часть клеток постоянно утрачивается, поэтому значительная часть клеток ткани находится в состоянии митоза (например, покровный эпителий, эпителий мочевыводящих путей).
В процессе митоза условно выделяют четыре стадии или фазы: профаза, метафаза, анафаза, телофаза.
Обязательным при митозе является построение митотического аппарата (ахроматиновое веретено). У животных клеток и клеток низших растений он образован системой микротрубочек и центриолями. У клеток высших растений веретено деления формируется при участии системы микротрубочек и сгустков цитоплазмы, формирующих у полюсов клетки - полярные шапочки. В ахроматиновом веретене образуются непрерывные нити, идущие от полюса к полюсу клетки и прерывистые, идущие к центру клетки от каждого из полюсов. Роль митотического веретена: обеспечивает направленное перемещение дочерних хромосом (хроматид) в анафазе; поддерживает форму клетки.
Основные события в фазах митоза.
Профаза (стадия материнского клубка). В этой фазе происходит распад ядерной оболочки, ядрышка, идет конденсация хромосом, слияние кариолимфы с гиалоплазмой, формируется веретено деления, прекращается синтез РНК (транскрипция). Хромосомы выглядят как клубок запутанных нитей, расщепляющихся с концов.
Метафаза (стадия "материнской звезды"). В этой фазе завершается формирование веретена деления (веретено состоит из непрерывных нитей, идущих от полюса к полюсу и прерывистых, идущих к центру клетки). Хромосомы максимально спирализованы, расщеплены на хроматиды, к центромерам таких хромосом (их кинетохорам) подходят прерывистые нити веретена деления и крепятся к ним, сокращаясь и растягиваясь нити "выстраивают" хромосомы на экваторе клетки перпендикулярно оси веретена деления. В этот период синтез белков практически не идет, клетки наиболее чувствительны к холоду, ядам. При действии средовых факторов (радиации, химических соединений, рентгеновского излучения) может произойти нарушение деления хромосом на хроматиды, что ведет к полиплоидии.
Анафаза (стадия "дочерних звезд"). Самая короткая фаза митоза, сокращение прерывистых нитей веретена деления ведет к разрыву центромеры и дочерние хроматиды оттягиваются за центромеры к противоположным полюсам клетки по непрерывным нитям веретена деления.
Телофаза ("профаза наоборот"). Процессы протекают порядке обратном профазе. Формируется ядерная оболочка. За счет ядрышкового организатора хромосом происходит формирование ядрышка. Нити веретена разрушаются, сосредоточившиеся на полюсах клетки хромосомы деспирализуются, удлиняются, в это время их невозможно увидеть в световой микроскоп.
Цитокинез - деление цитоплазмы и органоидов. Органоиды более или менее равномерно распределяются по двум полюсам. Мембрана животных клеток впячивается внутрь, в том месте, где был экватор веретена деления, образуется перетяжка, а затем выросты мембраны смыкаются, отделяя дочерние клетки друг от друга. Образование мембраны у животных клеток идет центростремительно. У растений на месте экватора выстраиваются пузырьки аппарата Гольджи и микротрубочки, образуя фрагмопласт. При слиянии содержимого пузырьков фрагмопласт превращается в серединную пластинку, общую для обеих клеток. Ее формирование идет центробежно.
Биологическое значение митоза заключается в строго одинаковом распределении реплицированного наследственного материала материнской клетки между дочерними, что обеспечивает образование генетически равноценных клеток и сохраняет приемственность в ряду поколений, обеспечивает рост. Следует отметить, что все процессы, происходящие в клетке, в том числе и митоз, находятся под генетическим контролем. Гены контролируют последовательные стадии редупликации ДНК, движение, цитокинез, спирализацию хромосом и т. д.
На основе митотического цикла возник ряд механизмов с помощью которых в том или ином организме количество наследственного материала может быть увеличено при сохранении постоянного числа клеток.
Эндомитоз или внутренний митоз. Практически происходит внутри ядра. При этом ядерная оболочка не разрушается и веретено не образуется, однако внутреннее содержимое ядра проходит все фазы характерные для митоза. В его основе лежит механизм удвоения ДНК, идентичный редупликации ДНК в митотическом цикле, но деление цитоплазмы нет (кратное увеличение числа хромосом). В результате получаются клетки с тетраплоидным (4п) или более высокой плоидности набором хромосом. Эндомитоз отмечен в клетках печени, клетках внутренней стенки пыльника, клетках антиподах.
Шизогония (множественное деление) характерна для некоторых простейших, ведущих паразитический образ жизни, когда одна материнская клетка образует до нескольких десятков дочерних. В случае шизогонии цитокинез запаздывает по сравнению с митозом, и сначала образуются ядра, а затем вокруг них формируются клетки. Это позволяет паразитическим простейшим быстро распространиться в теле организма-хозяина.
Политения Явление обнаружено в клетках слюнных желез насекомых, где найдены гиганские хромосомы. Политения роисходит за счет увеличения содержания ДНК в хромонемах хромосом. В таких клетках при сохранении диплоидного набора хромосом, количество генов повторено более, чем два раза, при этом увеличивается масса клетки, увеличиваются функциональные возможности организма.
Амитоз (прямое деление клеток) - разделение ядра путем перетяжки, без сложной перестройки наследственного материала. При этом типе деления клетка выигрывает в скорости, но проигрывает в "качестве", поскольку образовавшиеся дочерние клетки содержат наборы хромосом не идентичные материнскому. Таким способом делятся клетки, заканчивающие развитие или не нуждающиеся в полноценной генетической информации: клетки запасающих тканей семени - эндосперма, кожного эпителия, опухолевые клетки, стареющие клетки.
Мейоз
Мейоз (мeiosis с греческого - уменьшение). Мейоз - это деление созревания, особый способ деления клеток, в результате которого происходит редукция числа хромосом и переход клетки из 2п в п состояние. Мейоз - основное звено гаметогенеза. В. Флемминг 1882 году описал мейоз у животных, Э. Страсбургер 1888году наблюдал явление редукции числа хромосом у растений.
Мейоз может занимать различное положение в жизненном цикле. Для большинства животных характерна так называемая гаметическая редукция, в этом случае редукционное (мейотическое) деление непосредственно предшествует образованию гамет.
В зависимости от места в жизненном цикле организма различают 3 основных типа мейоза:
Зиготный (например, у грибов, водорослей). У некоторых простейших (например, у споровиков) редукция числа хромосом осуществляется сразу после образования зиготы и поэтому называется зиготической. При зиготической редукции, наоборот, преобладает гаплоидная фаза, а диплоидная представлена зиготой.
Гаметный - конечный (например, у многоклеточных животных, низших растений) идет в половых органах, где образуются гаметы. В случае гаметической редукции в жизненном цикле преобладает диплоидная стадия (гаплоидная представлена лишь гаметами).
Споровый - промежуточный (например, у высших растений) . Для всех высших растений характерна спорическая редукция, сопровождающая процесс спорогенеза (уменьшение числа хромосом происходит перед образованием спор, а не гамет). Этот тип редукции также называется смешанным, так как и диплоидная и гаплоидная фазы жизненного цикла существуют более или менее значительное время (при этом у моховидных преобладает гаплоидная стадия, называемая гаметофитом, а у всех других групп высших растений - диплоидная, называемая спорофитом).
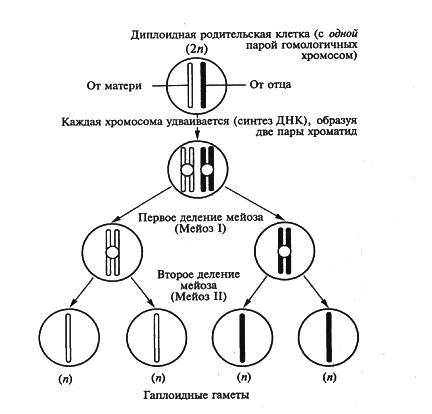
Мейоз включает 2 последовательных деления:
Мейотическое делении 1 или редукционное;
Мейотическое делении 2 или эквационное.
Между ними есть период покоя или интеркинез, однако редупликация ДНК происходит только перед первым делением. Как и митоз, каждое из двух мейотических делений разделяется на четыре фазы: профазу, метафазу, анафазу, телофазу.
В первом мейотическом делении профаза 1 самая продолжительная. Включает пять подстадий: лептотена, зиготена, пахитена, диплотена и диакинез.
Лептотена - начальная стадия профазы I, характеризуется тем, что хромосомы переходят в конденсированную форму и представлены в виде тонких длинных нитей, прикрепленных с помощью прикрепительного диска к ядерной мембране. При этом каждая хромосома состоит из двух сестринских хроматид, так как уже в интерфазе произошла репликация хромосомной ДНК, однако хроматиды тесно сближены и в световой микроскоп неразличимы.
Во время зиготены гомологичные хромосомы, представленные двумя хроматидами, сближаются и тесно примыкают друг к другу. Процесс сближения и "слипания" гомологичных хромосом называют конъюгацией (или синапсисом). В основе этого явления лежит специфическое сродство участков молекул ДНК. Кроме того, в зиготене осуществляется синтез небольшого количества ДНК, необходимой, по-видимому, для осуществления конъюгации.
Пахитена начинается тогда, когда синапсис хромосом полностью завершен. Сестринские хроматиды в каждой хромосоме становятся хорошо различимыми. Две полностью проконъюгировавшие гомологичные хромосомы образуют так называемый бивалент, а совокупность хроматид бивалента (их четыре - по две в каждой из гомологичных хромосом) - образуют тетраду. В поздней пахитене наблюдается формирование рекомбинационных узелков - структур, обеспечивающих рекомбинацию (обмен участками между гомологичными хромосомами).
В диплотене гомологичные хромосомы, составляющие бивалент, разъединяются. Но такое разделение обычно не бывает полным, н в местах расположения ре комбинационных узелков наблюдаются Х-образные соединения, называемые хиазмами, или перекрестами. В хиазмах обычно осуг ,ествляется обмен участков гомологичных хромосом - кросстговер. Он происходит благодаря разрывам и восстановлению нуклеотидной последовательности в молекуле ДНК. Благодаря тому, что отдельные участки молекулы ДНК (гены) несут информацию о формировании какого-либо признака, кроссинговер, обеспечивающий их обмен, представляет собой важнейший механизм наследственной изменчивости. На стадии диплотены хромосомы частично деконденсируются и становятся активными в отношении синтеза РНК. Коньюгация гомологичных хромосом и кроссинговер, дает материал для комбинативной изменчивости.
Во время диакинеза хромосомы вновь максимально конденсируются, отделяются от ядерной мембраны и располагаются по периферии ядра клетки, при этом очень четко различимы все 4 хроматиды бивалента. Каждая пара сестринских хроматид связана посредством центромер, а пара гомологичных хромосом - посредством хиазмов, которые возникают между двумя несестринскими хроматидами (хроматидами, принадлежащими отдельно каждой из двух спаренных хромосом).
Далее механизмы деления мало чем отличаются от обычных митотических.
В метафазе первого мейотического деления формируется метафазная пластинка и возникает веретено деления. Гомологичные хромосомы максимально спирализованы (биваленты выстраиваются в экваториальной плоскости), к их центромерам прикреплены нити веренена деления. Набор - 2п 4с.
В анафазе 1 - вследствие сокращения нитей веретена биваленты разрушаются (разрываются в местах хиазмов), к противоположным полюсам растаскиваются гомологичные хромосомы, состоящие из двух сестринских хроматид. Это ведет к редукции числа хромосом у каждого полюса. Набор у каждого полюса становится п2с.
Телофаза 1 и интеркинез у большинства имеется, но не всегда обязательны. Телофаза I очень короткая. Она характеризуется формированием новых ядер и ядерной мембраны. К концу телофазы 1 формируются 2 клетки с набором п 2с. Разделения хроматид, соединенных в области центромеры, не происходит.
За телофазой следует обычно очень непродолжительная интерфаза второго мейотического деления (интеркинез), причем для нее характерно отсутствие синтетического (S) периода. Таким образом, перед вторым мейотическим делением репликации ДНК не происходит и диплоидность не восстанавливается.
Второе мейотическое (эквационное) деление протекает по принципу митоза:
первые две фазы иногда выпадают;
в профазе 2 ядерные оболочки разрушаются, центриоли (в клетках животных) перемещаются к противоположным полюсам;
в метафазе формируется метафазная пластинка и веретено деления;
в анафазе 2 расходятся хроматиды (продольные половинки) оставшихся хромосом, что сохраняет гаплоидность и уменьшает наполовину наследственную информацию в каждой из образующихся клеток (пс); хроматиды перемещаются к противоположным полюсам;
в телофазе 2 исчезают нити веретена деления и происходит обособление ядра, происходит цитокинезом. Заканчивается весь процесс мейоза с образованием 4 гаплоидных клеток с набором хромосом (п) и с половинной наследственной информацией в них (с).
Таким образом, в результате двух последовательных мейотических делений каждая исходно диплоидная клетка дает начало четырем гаплоидным дочерним клеткам, генетически разнородным с гаплоидным набором (п), при слиянии которых при оплодотворении происходит восстановление диплоидности в зиготе, содержащей полную наследственную информацию, характерную для данного вида.
Мейоз создает возможность для возникновения в гаметах новых генных комбинаций, что ведет к изменениям в генотипе и фенотипе потомства.