|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Полиморфизм ДНК Y-хромосомы.
Генетическая история народов Волго-Уральского региона по данным полиморфизма мт ДНК и У хромосомы. В Волго-Уральском регионе столкнулись две волны расселения: европеоидная и монголоидная. Находясь на границе двух частей света - Европы и Азии, этот регион на протяжении исторически длительного времени был местом взаимодействия многих этнических слоев. Современные популяции Волго-Урала крайне неоднородны по этнолингвистической структуре. Сейчас здесь проживают представители финно-угорской ветви Уральской языковой семьи (удмурты, марийцы, мордва, коми), тюркской ветви Алтайской языковой семьи (башкиры, татары, чуваши) и восточно-славянской ветви Индоевропейской языковой семьи (русские). С целью познания факторов формирования генофонда, генетической реконструкции родства и происхождения современных народов Волго-Урала изучены 17 полиморфных аутосомных ДНК-локусов, определена нуклеотидная последовательность ДНК гипервариабельного участка митохондриального генома, выполнен анализ полиморфизма 24 локусов Y-хромосомы. В ходе исследований проанализированы геномы 1500 индивидов из 8 популяций (башкиры, татары, русские, чуваши, удмурты, коми, мордва, марийцы).
Полиморфизм мтДНК. Чтобы судить о генетическом разнообразии народов Волго-Уральского региона по материнской линии, была определена последовательность гипервариабельного участка митохондриальной ДНК (377 нуклеотидов) и проведен анализ полиморфизма мтДНК с использованием 26 эндонуклеаз (ферментов рестрикции). Полученные данные анализировались методом так называемых медианных сетей. Медианная сеть - один из графических вариантов представления данных о полиморфизме ДНК, который позволяет проследить дивергенцию митохондриального генома от предковой ДНК. В качестве предкового выбирается гаплотип, который широко распространен в данной популяции и включает наиболее частые аллели, характерные для данного этноса. В процессе такого анализа в единую медианную сеть соединяются посредством линий сотни и тысячи соседствующих, наиболее близких индивидуумов. В этой сети узловые соединения совпадают со специфическими гаплотипами, определенными по характерным мутациям изучаемого генома. Для большинства расовых и географических групп установлена специфичность типов мтДНК. Более 90% всех типов мтДНК народов Западной Евразии принадлежит к линиям, которые рассматриваются как потомки европейского генного пула, существовавшего в верхнем палеолите. Митохондриальный геном азиатских популяций включает два суперкластера, которые соединяются в африканском макрокластере - общий предок для всех неафриканских популяций мира. Около 50% линий мтДНК коренных жителей Азии принадлежит к суперкластеру, который подразделяется на линии.
Большинство типов мтДНК народов Волго-Уральского региона соответствует линиям мтДНК Европы и Ближнего Востока, что свидетельствует об общих предковых линиях мтДНК, специфичных для европейцев. Анализ европеоидного и монголоидного вклада в материнские генетические линии народов Волго-Уральского региона не выявил корреляции языка и геномного состава этнических групп. Сравнительный анализ типов мтДНК в 18 популяциях Евразии, включая популяции Волго-Уральского региона позволил установить западно-восточный градиент увеличения частоты азиатских линий мтДНК. Установлено, что лингвистическое сходство популяций играет меньшую роль, чем географическая близость или отдаленность популяций. Один из наиболее важных аспектов анализа митохондриального генома - оценка времени коалесценции (расхождения, дивергенции) линий мтДНК в пределах каждой линии. Безусловно, на временные оценки будут влиять различные факторы формирования разнообразия мтДНК: объем выборки, миграция населения, резкий рост численности, феномен "бутылочного горлышка" - сильное сокращение численности наших предков, вызванное, по-видимому, изменением климата, и т.д. Тем не менее оценка времени дивергенции линии возможна при обнаружении предковых гаплотипов.
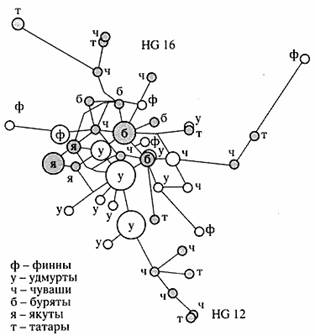
Полиморфизм ДНК Y-хромосомы. Генетические свойства Y-хромосомы, такие как передача только по отцовской линии, отсутствие рекомбинации, малая эффективная численность пула Y-хромосом по сравнению с аутосомами (в четыре раза меньше, чем у аутосом), позволяют прослеживать по гаплотипам Y-хромосомы отцовские линии, представляющие собой последовательную "запись" мутаций в ряду поколений. По сравнению с митохондриальным геномом, насчитывающим 16.5 тыс. пар нуклеотидов. Y-хромосома, размер которой оценивается приблизительно в 60 млн. пар нуклеотидов, дает в руки исследователей потенциально более мощное "оружие". Для изучения и сравнения генетического разнообразия отцовских линий в популяциях Волго-Уральского региона использовались 24 маркера Y-хромосомы. В качестве примера на рисунке 5 представлена медианная сеть линий 12 и 16 Y-хромосомы как наиболее интересных в контексте финноугорских народов. Линия 16 практически отсутствует в западноевропейских популяциях.
Для линии 16 Y-хромосомы уровень генетического разнообразия намного выше в популяциях Восточной Европы (чуваши, татары). Анализ распространения и разнообразия линии 16 Y-хромосомы среди восточноевропейских популяций показывает, что местом ее "рождения", возможно, является Восточно-Европейская равнина. Согласно филогеографическому анализу этой линии в Евразии, она начала распространяться с запада на восток. В то же время частота распространения линии 12 - предковой для линии 16 - меньше, чем для линии 16. Особенно характерна для популяций Волго-Уральского региона линия 3 Y-хромосомы, частота встречаемости которой максимальна у славян (русские и поляки), а также у населения Латвии, Литвы и Эстонии. Судя по результатам анализа отцовских линий в популяциях Волго-Уральского региона, главную роль в формировании генетического разнообразия народов, проживающих на этой территории, по-видимому, играет географическая близость, а не языковая принадлежность. Исследования полиморфизма аутосомных, митохондриальных и Y-хромосомных ДНК-маркеров внесли важный вклад в понимание путей происхождения человека и рас, расселения Homo sapiens по планете, в генетическую и демографическую историю отдельных этносов и популяций.
Поиск по сайту: |