|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Регистрация и преобразование обонятельного сигнала ⇐ ПредыдущаяСтр 2 из 2
Рецепторные клетки обонятельной выстилки регистрируют небольшое количество первичных запахов, но их комбинации формируют ощущение многих и многих воспринимаемых запахов. Возникает вопрос: каким образом обонятельная система может различать множество различных запахов? Варианты ответа таковы: или рецепторные обонятельные нейроны имеют множество различных молекулярных рецепторов (в том числе и каждый нейрон), или/и содержат один или несколько типов молекулярных обонятельных рецепторов, но посылают в ЦНС различные спектры ПД. Наконец, формирование ощущения конкретного запаха может зависеть от фиксированных связей между нервными клетками обонятельной системы.
Обонятельные рецепторные белки кодируют гены, расположенные практически во всех хромосомах, кроме аутосомы 20 и половой хромосомы Y. В геноме человека идентифицировано более 900 генов, кодирующих белки обонятельных рецепторов, что практически равно одной тридцатой всего генома. Эти белки относят к семейству связанных с G‑белком рецепторов. Последовательность событий при регистрации обонятельного сигнала (рис. 12–5) может быть представлена следующим образом: взаимодействие пахучего вещества с белком-рецептором в плазмолемме обонятельных волосков ® активация G‑белка ® повышение активности аденилатциклазы ® увеличение уровня цАМФ ® активация цАМФ–зависимых воротных катионных каналов ® деполяризация рецепторных нейронов ® генерация ПД и его проведение по аксону.
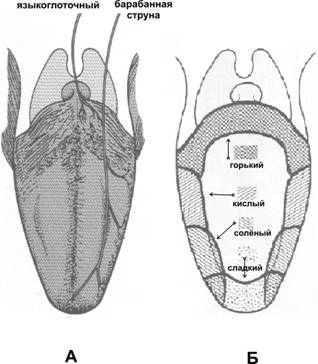
Рис. 12–5. Трансформация сигнала в обонятельном рецепторе. А — рецепторный обонятельный нейрон; Б — вход Na+ в клетку; В — пахучие молекулы соединяются с рецептором (R). Рецептор активирует G‑белок (G), G‑белок активирует аденилатциклазу (Ац), образующийся цАМФ открывает Na+‑каналы. Система инозитолтрифосфата (ИТФ) также имеет отношение к механизму хемовосприятия в органе обоняния. При действии некоторых пахучих веществ быстро возрастает уровень ИТФ, который взаимодействует с Ca2+‑каналом в плазмолемме обонятельных рецепторных нейронов. Таким образом, системы вторых посредников цАМФ и ИТФ взаимодействуют между собой, обеспечивая лучшее различение запахов. Механизмы адаптации. Адаптация к обонятельному сигналу на уровне рецепторных нейронов (регуляция чувствительности к обонятельному сигналу) происходит достаточно быстро (50% в течение 1 с). Некоторые механизмы такой быстрой адаптации известны и осуществляются при помощи внутриклеточных вторых посредников и аррестинов. · При действии некоторых пахучих веществ в обонятельных рецепторных нейронах быстро увеличивается содержание инозитолтрифосфата, взаимодействующего с Ca2+‑каналом. Через цАМФ–зависимые воротные ионные каналы внутрь клетки проходят не только одновалентные катионы, но и Ca2+, связывающийся с кальмодулином. Образовавшийся комплекс Ca2+-кальмодулин взаимодействует с каналом, что препятствует его активации при помощи цАМФ, в результате чего рецепторная клетка становится нечувствительной (адаптируется) к действию пахучего вещества. · Обонятельные реснички содержат множество молекул аррестинов. Эти белки, связанные с b‑адренорецепторами и G‑белками, участвуют в десенситизации обонятельных рецепторов. Центральные механизмы Пребывание в условиях воздействия даже очень неприятного запаха уменьшает и может полностью подавить восприятие запаха. Этот феномен — результат быстрой адаптации или десенситизации на уровне рецепторных нейронов (см. выше). При этом порог для других запахов не изменяется. В центральных обонятельных структурах также существует чёткий контроль обонятельной афферентной импульсации. Так, возбуждение нервных клеток, вызванное афферентной импульсацией, сменяется последующим торможением электрической активности нейронов, что и объясняет существование ритмической активности в обонятельной коре. Пример центральной регуляции потоков импульсов от обонятельной луковицы приведён выше. Обонятельная импульсация направляется в лимбическую систему (здесь осуществляется подсознательное восприятие обоняния) к передним обонятельным ядрам, миндалевидному телу (формирование эмоционального ответа на обонятельные стимулы) и обонятельной борозде (формирование так называемой обонятельной памяти). Обоняние, половое поведение и память. Между запахом и половой функцией у многих видов животных существует тесная взаимосвязь (скорее всего, реализуемая через cошниково–носовой [вомероназальный] орган Якобсона — участок слизистой оболочки носовых ходов, аналогичный обонятельной выстилке), а использование духов даёт достаточные основания считать, что подобная взаимосвязь существует и у людей (орган Якобсона у человека отсутствует). Обоняние у женщин развито сильнее, и оно обостряется ещё больше в период овуляции. Запах и (в меньшей степени) вкус обладают уникальной способностью оживлять воспоминания, заложенные в долговременной памяти. Этот факт отмечен писателями и экспериментально доказан психологами. Комбинаторное кодирование. Эта концепция подразумевает, что каждый из миллионов отдельных запахов (одорантов) имеет уникальный код; кодирование происходит на уровне органа обоняния, а декодирование — в обонятельных центрах. В пользу существования такого обонятельного кода свидетельствуют следующие факты. · Каждый обонятельный нейрон экспрессирует один тип обонятельных рецепторов. · Конкретный тип обонятельного рецептора распознаёт несколько одорантов (в среднем четыре). · Конкретная молекула одоранта может активировать несколько разных типов обонятельных рецепторов. · Минимальные изменения конформации одоранта, его химической структуры, а также изменения концентрация молекулы пахучего вещества изменяют код одоранта. Последнее обстоятельство позволило объяснить феномен формирования ощущения разных запахов при вдыхании разных концентраций одного и того же одоранта. Нарушения обоняния включают аносмию (отсутствие чувства запаха), гипосмию (уменьшение обонятельной чувствительности), дизосмию (искажение чувства обоняния), гиперосмию (усиление обоняния). У человека известно несколько видов аносмий, касающихся отсутствия восприятия запаха для какого-нибудь одного из членов семейства обонятельных рецепторов. Пороги восприятия запахов увеличиваются с возрастом. Примерно 75% лиц старше 80 лет имеют нарушенное восприятие запахов. Вкус Рецепторы вкуса — контактные, а пути проведения от них проходят через ствол мозга к таламусу и проецируются вдоль постцентральной извилины. Периферическая часть вкусового анализатора — вкусовые почки — расположены в слизистой оболочке полости рта, переднего отдела глотки, пищевода и гортани. Основная масса вкусовых почек (>90% их общего количества — до 10 тыс.) находится в хемочувствительных сосочках языка (рис. 12–6) — листовидных, грибовидных и желобоватых.
Рис. 12–6. Вкусовые области языка. Вкус сладкого регистрируется преимущественно на кончике языка, солёного — ближе к кончику языка, кислого — на боковых сторонах языка, горького — в задней части языка и в мягком нёбе. · Листовидные сосочки развиты у детей (у взрослых они атрофированы), образуют две группы (4–8 сосочков в каждой) по левому и правому краям тела языка. · Грибовидные сосочки (около 100) расположены по дорсальной поверхности передних двух третях языка, рисунок их расположения индивидуален. · Желобоватые сосочки (6–12 штук) расположены в задней части языка, кпереди от пограничной борозды между телом и корнем языка. Вкусовые почки (вкусовые луковицы) — тельца яйцевидной формы со средним размером 50–70 мкм, расположены в пласте многослойного плоского эпителия, занимая всю толщу пласта, от базальной мембраны эпителия до его поверхности. Все клетки вкусовой луковицы постоянно обновляются за счёт окружающих клеток эпителия, но только при условии интактной чувствительной иннервации той области слизистой оболочки, где расположены вкусовые луковицы. Время жизни клеток вкусовой луковицы составляет 10 сут. Каждая вкусовая почка содержит поддерживающие (опорные) и вкусовые (рецепторные) клетки (рис. 12–7).
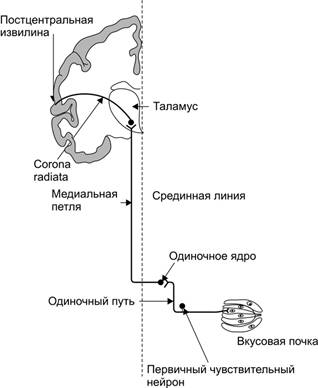
Рис. 12–7. Вкусовая почка состоит из удлинённых светлых клеток различных типов. На верхушечной (апикальной) части большинства клеток расположены микроворсинки — вкусовые волоски, занимающие вкусовой канал, открывающийся на поверхность эпителия вкусовой порой. В микроворсинки «вмонтированы» рецепторные вкусовые белки. В базальной части вкусовой луковицы рецепторные клетки образуют синапсы с терминальными разветвлениями периферических отростков первичных чувствительных нейронов. Каждая вкусовая почка иннервирована примерно 50 нервными волокнами, а каждый чувствительный нейрон получает сигналы примерно от 5 вкусовых луковиц. [11]. Регенерация вкусовых клеток. Во вкусовом рецепторе происходит постоянное обновление клеток. Из периферической области вкусовой почки клетки перемещаются в центральную её часть со скоростью 0,06 мкм/час. Средняя продолжительность жизни клеток вкусового рецепторного органа составляет 250±50 часов. После повреждения нервов, иннервирующих вкусовые почки, последние дегенерируют, а при регенерации нервов происходит их восстановление. Восходящие пути вкусовой чувствительности состоят из трёх нейронов: 1. первичного чувствительного нейрона; 2. нервной клетки ядра одиночного пути (одиночное ядро, nucleus tractus solitarius); 3. нейрона вентромедиального ядра таламуса (рис. 12–8).
Рис. 12–8. Восходящие пути вкусовой чувствительности. Часть аксонов одиночного пути направляется к верхнему и нижнему слюнным ядрам, что обеспечивает выполнение некоторых рефлексов, связанных со слюноотделением. 1. Первичные чувствительные нейроны · Чувствительные нервные волокна от вкусовых почек, расположенных в передних двух третях языка (рис. 12–6А), проходят в составе язычного нерва и далее — барабанной струны (chorda tympani системы лицевого нерва). Тела первичных чувствительных нейронов находятся в узле коленца (ganglion geniculi). Центральные отростки этих нейронов в составе промежуточного нерва (n. intermedius) и одиночного пути подходят к одиночному ядру (nucleus tractus solitarius). · Афферентные волокна от вкусовых луковиц задней трети языка (рис. 12–6А) проходят в составе языкоглоточного нерва. Тела первичных чувствительных нейронов находятся в нижнем узле (ganglion inferius), расположенном у выхода из яремного отверстия. Центральные отростки нейронов в составе одиночного пути направляются к одиночному ядру. · Чувствительные нервные волокна из более каудально расположенных областей входят в состав блуждающего нерва. Тела первичных чувствительных нейронов находятся в верхнем и нижнем узлах (ganglii superius et inferius), а их центральные отростки в составе одиночного пути направляются к одиночному ядру. 2. Нейроны ядра одиночного пути. Вторые нейроны восходящих путей вкусовой чувствительности расположены в ростральной части ядра восходящего пути. Входящие в ядро центральные отростки первичных чувствительных нейронов образуют синапсы с дендритами нервных клеток ядра, а их аксоны переходят на другую сторону, присоединяются к медиальной петле (lemniscus medialis) и следуют до вентромедиального ядра таламуса. 3. Нейроны вентромедиального ядра таламуса. Аксоны нервных клеток ядра таламуса в составе лучистого венца (corona radiata) направляются к вентральной части постцентральной извилины. Представительство вкуса не имеет отдельной корковой проекционной области, оно представлено в участке постцентральной извилины, получающем проекции кожной чувствительности лица. Основные вкусы. Человек различает четыре первичных вкуса (сладкий, кислый, горький и солёный), а также «умами» (от японского «изысканный», вкус глутамата натрия). Существуют карты топографической привязанности преимущественного восприятия основных вкусов (см. рис. 12–6А), но необходимо иметь в виду, что такие карты справедливы только для низких концентраций тестируемых вкусовых веществ. Так, при низких концентрациях деполяризуются и формируют рецепторный (генераторный) потенциал лишь отдельные вкусовые клетки. Некоторые вкусовые клетки и вкусовые луковицы отвечают преимущественно на горькие стимулы, в то время как другие — на сладкие, кислые или солёные. Часть вкусовых клеток и луковицы в целом отвечают на две или три вкусовые модальности, а иные даже на все. При увеличении же концентрации вкусовых веществ происходит возбуждение практически во всех вкусовых луковицах самой различной локализации. Вкусовой порог. Наиболее значительные различия наблюдают между веществами, создающими ощущение горького и всех остальных первичных вкусов. Так, ощущение солёного возникает при воздействии вещества в концентрации 0,01 М, а наличие хинина можно выявить при его концентрации в миллион раз меньшей. Пороги вкусовой чувствительности у разных лиц заметно различаются. Значительное влияние на чувствительность оказывает состояние организма (например, стресс, беременность, голодание). Часто наблюдают «вкусовую слепоту» (классический пример — производные тиомочевины, например фенилтиокарбамид, присутствие которого не чувствует каждый пятый). Способность человека различать интенсивность вкусовых ощущений сравнительно груба. Так, 20% раствор сахара ощущается как максимально сладкий, 10% раствор поваренной соли как максимально солёный и т.д.
Адаптация к воздействию вкусового вещества развивается медленно (минуты) и пропорциональна его концентрации. К сладкому и солёному адаптация развивается быстрее, чем к горькому и кислому. Нарушения вкуса. К нарушениям вкуса относят агевзию (отсутствие чувства вкуса), гипогевзию (уменьшение вкусовой чувствительности) и дисгевзию (извращение вкуса). Гипогевзию наблюдают при многих заболеваниях. Некоторые вещества (например, содержащие сульфгидрильные группы каптоприл и пеницилламин) вызывают временную потерю вкусовой чувствительности.
Поиск по сайту: |