|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Функции плазмалеммы, общие для плазматических мембран.
ПЛАЗМАТИЧЕСКАЯ МЕМБРАНА РАСТИТЕЛЬНОЙ КЛЕТКИ (ПЛАЗМАЛЕММА)
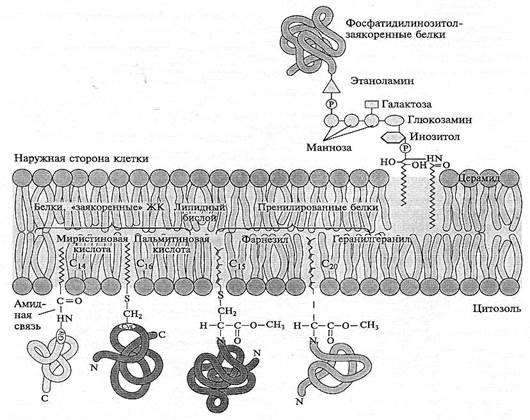
СТРУКТУРА ПЛАЗМАЛЕММЫ Общая характеристика.Плазматическая (цитоплазматическая) мембрана — обязательный компонент любой клетки. Она отграничивает клетку и обеспечивает сохранение существующих различий между клеточным содержимым и окружающей средой. Мембрана служит высокоизбирательным «входным» селективным фильтром и отвечает за активный транспорт веществ в клетку и из нее. Цитоплазматическую мембрану растительной клетки обычно называют плазмалеммой. Как и любая биологическая мембрана, она представляет собой липидный бислой с большим количеством белков. Основу липидного бислоя составляют фосфслипиды. Помимо них в состав липидного слоя входят гликоли-пиды и стерины. Липиды достаточно активно перемещаются в пределах своего монослоя, но возможны и их переходы из одного монослоя в другой. Такой переход, называемый «флип-флоп» (от англ. flip-flop), осуществляется ферментом флипазой. Кроме липидов и белков в плазмалемме присутствуют углеводы. Соотношение липидов, белков и углеводов в плазматической мембране растительной клетки составляет приблизительно 40 :40 : 20. Мембранные белки связаны с липидным бислоем различными способами. Первоначально белки мембран разделяли на два основных типа: периферийные и интегральные. Периферийные белки ассоциированы с мембраной за счет присоединения к интегральным белкам или липидному бислою слабыми связями: водородными, электростатическими, солевыми мостиками. Они в основном растворимы в воде и легко отделяются от мембраны без ее разрушения. Некоторые периферийные белки обеспечивают связь между мембранами и цитоскелетом. Интегральные белки мембран нерастворимы в воде. Как минимум один из доменов интегрального белка встроен в гидрофобную часть бислоя мембраны, поэтому интегральный белок, как правило, не может быть удален из мембраны без ее разрушения. В последнее время показано существование третьей группы белков, так называемых «заякоренных» в мембране белков (рис. 1.4). Эти белки фиксируются в мембране за счет специальной молекулы, в качестве которой могут выступать жирная кислота (ЖК), стерин, изопреноид или фосфатидилинозитол.
Рис. 1.4. Варианты фиксации «заякоренных» в мембранах белков
Белки, связанные с изопреноидами (пренилированные белки) или жирной кислотой, обратимо соединяются с эндоплазматической (внутренней) поверхностью мембраны. Из жирных кислот чаще используется миристиновая (С14) или пальмитиновая (С16). В первом случае образуется амидная связь с терминальной аминогруппой глицина. К остаткам пальмитиновой кислоты белки присоединяются за счет тиоэфирных связей с цистеинами в С-конце полипептидной цепи. Для пренилирования белков обычно используется фарнезил или геранилгеранил, которые также присоединяются к остаткам цистеина в карбоксильном конце полипептида. В отличие от этих двух групп белков фосфатидилинозитолсвязанные белки находятся с внешней (экстрацеллюлярной, или люменальной) стороны мембраны. Подобным образом, по-видимому, связаны с плазматической мембраной большинство арабиногалактановых белков. Холестеринсвязанные белки недавно были обнаружены в плазматической мембране животных клеток, но в растительных клетках подобные белки пока не найдены. Особенностью липидного состава плазмалеммы по сравнению с другими мембранами растительной клетки является высокое содержание стеринов, но в отличие от плазматической мембраны животной клетки для плазмалеммы характерна высокая вариабельность их состава в зависимости от вида растения, органа и ткани. Например, у ячменя {Hordeum vulgaris) в клетках корня количество свободных стеринов превышает количество фосфслипидов более чем в два раза, тогда как в листьях фосфслипидов больше, чем стеринов почти в 1,5 раза. В листьях шпината (Spinacia oleracia) соотношение фосфслипиды: свободные стерины почти на порядок выше — 9:1. Структурные особенности плазмалеммы.Помимо высокой степени вариабельности плазматическая мембрана растительной клетки имеет ряд структурных особенностей, отличающих ее от других эукариотических клеток. • Жирнокислотный состав. Основными жирными кислотами плазмалеммы являются пальмитиновая (16:0), олеиновая (18 :1; Δ9), линолевая (18:2; Δ9,12) и линоленовая (18:3; Δ9,12 ‘15). Практически отсутствует стеариновая кислота (18:0) и полностью — арахидоновая (20 :4; Δ5’ • Набор стероидов. В плазмалемме, как и во всех мембранах растительной клетки, почти нет холестерина. Его заменяют фитостерины, которые являются С-24-замещенными стеринами (рис. 1.5). Основные фитостерины — ситостерин, стигмастерин и кампестерин. Помимо свободных фитостеринов присутствуют значительные количества эфиров, гликозидов и ацилгликозидов стеринов, что не характерно для клеток животных. • Набор белков. Наличие специфических белков, прежде всего арабиногалактанов (AGPs), обеспечивающих структурное и функциональное взаимодействие плазмалеммы с клеточной стенкой.
Рис. 1.5. Структура фосфолипидов (А) и фитостеринов (Б)
ФУНКЦИИ ПЛАЗМАЛЕММЫ Плазмалемма, будучи плазматической мембраной, выполняет все общие для этих структур функции, однако некоторые из них она выполняет особым образом. Кроме того, плазмалемма имеет ряд специфичных функций, характерных только для растительной клетки (например, синтез целлюлозы, осуществление контакта с клеточной стенкой). Функции плазмалеммы, общие для плазматических мембран. 1. Контроль поглощения и секреции веществ. Вплазмалемме находятся множество транспортных систем, прежде всего для транспорта ионов, — ионные каналы, ионные переносчики и ионные насосы. Благодаря им осуществляется очень точный и селективный транспорт ионов как внутрь клетки, так и наружу, т.е. оптимальное снабжение клетки необходимыми ионами (подробнее о механизмах этого процесса см. в гл. 6 «Минеральное питание»). Через плазмалемму происходит также перенос макромолекул. Так, в периплазматическое пространство транспортируются строительные блоки клеточной стенки — полисахариды и структурные белки. Эти соединения обычно находятся в везикулах аппарата Гольджи и высвобождаются из них путем экзоцитоза. 2. Запасание и использование энергии. Плазмалемма любой клетки является энергизованной мембраной, т.е. на ней существует градиент электрохимического потенциала Δ 3. Размещение и обеспечение работы ферментов.Вплазмалемме находится множество ферментов. Это ферменты построения клеточной стенки (целлюлозосинтаза), ферменты сигнальных систем (фосфолипазы С, А; аденилатциклаза) и ряд других ферментов (например, цитохром Ь5, возможно — десатуразы). 4. Рецепторные функции.Многие белки плазмалеммы являются рецепторами различных сигналов. Можно сказать, что плазмалемма клетки — это мозаика различных рецепторов эндогенных сигналов (прежде всего фитогормонов) и внешних воздействий. К последним относятся рецепторы элиситоров (веществ, выделяемых патогенами) и рецепторы физических факторов — температуры, давления и др. Температура изменяет жесткость (текучесть) мембраны, т.е. ее механические свойства. Изменение этих свойств плазмалеммы приводит к открытию или закрытию находящихся в ней механосенсорных ионных каналов. Таким образом, плазмалемма является местом рецепции (восприятия) различных сигналов химической и физической природы. 5. Сигнальная функция.Многие компоненты плазмалеммы после восприятия сигналов служат источником вторичных мессенджеров — молекул, которые «передают» сигнал по эстафете и усиливают его. В качестве вторичных мессенджеров выступают инозитол-1,4,5-трифосфат, диацилглицерин, фосфатидная кислота, продукты перекисного окисления липидов мембран. Все эти соединения образуются из липидов плазмалеммы под действием специальных ферментов, активируемых в результате воздействия сигнала на рецепторы. Таким образом, плазматическая мембрана является местом не только рецепции сигналов, но также их усиления и дифференцирования (см. гл. 7).
Поиск по сайту: |
 ); характерные для мембран клеток животных и грибов. Известно, что арахидоновая кислота в очень низких концентрациях является мощным стимулятором фитоиммунитета для некоторых видов растений (например, обеспечивает устойчивость картофеля к фитофторе).
); характерные для мембран клеток животных и грибов. Известно, что арахидоновая кислота в очень низких концентрациях является мощным стимулятором фитоиммунитета для некоторых видов растений (например, обеспечивает устойчивость картофеля к фитофторе).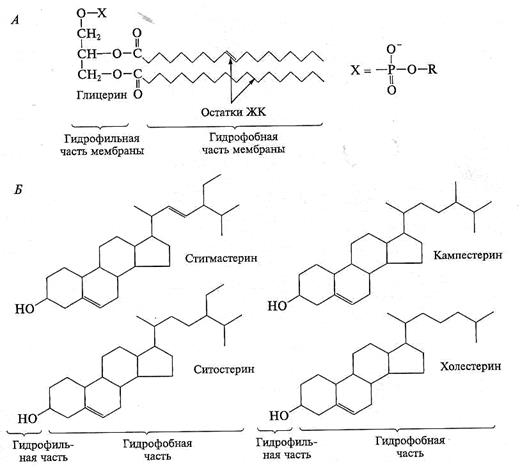
 , который используется для выполнения полезной работы, прежде всего для активного переноса веществ через мембрану (подробнее о механизмах см. в гл. 4 и 6).
, который используется для выполнения полезной работы, прежде всего для активного переноса веществ через мембрану (подробнее о механизмах см. в гл. 4 и 6).