|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Одномембранні компартменти.Стр 1 из 3Следующая ⇒
Внутрішні мембрани клітини.
Клітина живих організмів – складна багатофункціональна система. Відповідно і будова її надзвичайно складна. Під плазматичною мембраною розташовується вміст клітини, який носить назву протопласту. В клітинах еукаріот весь вміст клітини, але без ядра називають цитоплазмою. Якщо вилучити з цитоплазми всі структури, то залишиться рідина з розчиненими в ній органічними та неорганічними сполуками – гіалоплазма. Гіалоплазма є рідким внутрішнім середовищем клітини, що забезпечує обмін речовин та енергетичні перетворення. Часто гіалоплазму ще називають цитозолем. В цитології можна зустріти такі поняття як ектоплазма, яким позначають шар цитоплазми, що прилягає до плазматичної мембрани та ендоплазма – внутрішній шар. Цими поняттями позначають цитоплазму тваринних клітин. Але основою метаболічних процесів є унікальна властивість клітинних мембран – здатність до вибіркового транспорту речовин. Тому можна передбачити, що і вміст клітини буде також поділений на окремі відсіки, оскільки в ній одночасно протікають різноманітні біохімічні процеси. Для кожної групи хімічних реакцій необхідні свої специфічні умови та набір реагентів. Такі спеціальні умови можливо створити лише використавши мембранні структури. Дійсно, в цитоплазмі розташовуються окремі ділянки оточені внутрішніми мембранами. Вони отримали назву компартменти (від чеської – маленька квартирка), або відсік. Більш традиційна назва компартментів – органоїди або органели, хоча ці назви більш широкі і об‘єднують також і немембранні компоненти, наприклад, рибосоми. Кожний компартмент являє собою окрему герметично ізольовану комірку із власним специфічним набором ферментів. За фізичними та хімічними властивостями внутрішні мембрани відрізняються від плазматичної і одна від одної. До складу еукаріотичної клітини входять сім основних компартментів. Це ядро, де синтезується ДНК та РНК, ендоплазматичний ретикулум, апарат Гольджі, лізосоми, пероксисоми, мітохондрії та пластиди. З них оточені лише однією мембраною – ендоплазматичний ретикулум, комплекс Гольджі, лізосоми, пероксисоми та подвійною мембраною – ядро, мітохондрії і хлоропласти. Крім цих структур розрізнюють не мембранні компоненти – рибосоми, клітинний центр, клітинна стінка, глікокалікс, мікротрубочки, мікрофіламенти, клітинні включення. В структурі клітини мембрани утворюють певну систему. Так плазматична мембрана контактує з системою мембран ендоплазматичного ретикулуму, а той тісно поєднаний з комплексом Гольджі та лізосомами. Розглянемо загальну будову та функції окремих компартментів.
Одномембранні компартменти.
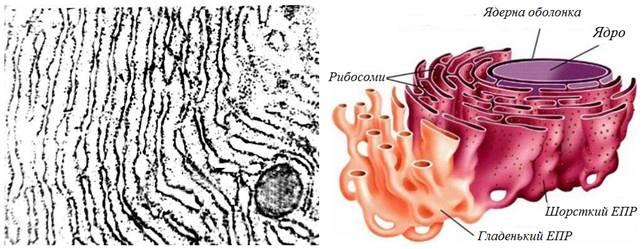
Ендоплазматичний ретикулум(скорочено ЕПР).Походження назви цього компартменту досить просте. Endos – внутрішній; plasma – утвір або виліплений – це стара і не зовсім вірна назва протоплазми; reticulum – сіточка. Дещо раніше цей компартмент носив назву ендоплазматичної сітки. Вважається, що ЕПР хоч і має чисельні складки та вигини, але являє собою єдину замкнену порожнину. Ця порожнина складає до 10% загального об’єму клітини. На електронних мікрофотографіях ЕПР добре помітні дві окремі ділянки. Це шорсткий або гранулярний ЕПР та гладенький або агранулярний. Гранулярний ЕПР містить рибосоми, які прикріплені до мембрани з боку, що повернутий до цитоплазми. Агранулярний ЕПР не має рибосом і являє собою розгалужену частину самої мембрани. За формою ці дві ділянки також відрізняються. Гранулярний утворений мішечками, що укладені у вигляді стопки. Такі мішечки отримали назву цистерни. Гладенький ЕПР представлений сіткою тонких трубочок. Гранулярний ЕПР тісно пов’язаний із зовнішньою мембраною ядра клітини і складає з ним одне ціле (рис. 37). Було встановлено, що на гранулярний ЕПР багаті клітини, які активно синтезують білок. Це дозволило припустити, а пізніше і експериментально довести, що саме на цьому компартменті синтезуються білкові молекули та молекули ліпідів. Навпаки, агранулярний ЕПР не бере участі в такому синтезі.
Рис. 37. Схематична будова ендоплазматичного ретикулуму (ЕПР). зліва – фрагмент ЕПР під електронним мікроскопом; зправа – тримірна модель шорсткої та гладенької ЕПР.
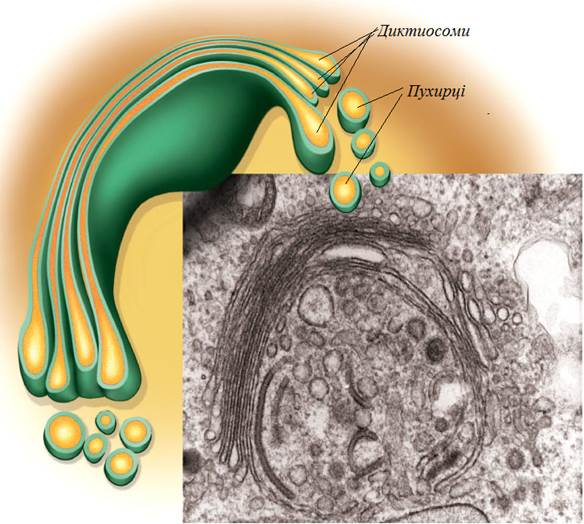
Це ділянки, що вільні від рибосом, є спеціалізованими зонами, де утворюються так звані транспортні пухирці. Такі пухирці оточені мембраною, яка відбрунькувалася від ЕПР. Вони упаковують новоутворені молекули білку, а також ліпіди, які транспортуються в різні ділянки клітини. Крім того гладенький ЕПР бере активну участь у нейтралізації та виведенні метаболітів, в тому числі і лікарських препаратів. Наприклад, в клітинах печінки при надходженні до них значної кількості фенобарбіталу (снодійного препарату) загальна поверхня агранулярного ЕПР за кілька днів збільшується вдвічі. Після виведення цього препарату через п’ять днів загальна площа цієї ділянки ЕПР нормалізується. Таким чином ЕПР є своєрідною фабрикою синтезу білків і ліпідів. Його порожнина має набір багатьох ферментів, що забезпечують такий синтез. Новоутворені органічні молекули спочатку надходять до ЕПР, а потім транспортуються до інших органоїдів. Вважають, що мембрани всіх інших компартментів побудовані з фосфоліпідів і білків, які синтезувалися на ЕПР. Апарат (комплекс) Гольджі.Вперше цей компартмент описав італієць Камілло Гольджі в 1898 році. Звичайно апарат Гольджі розташований поблизу ядра клітини. Він представлений системою гладеньких мембран, які утворюють сплощені дископодібні цистерни. Цистерни складені у вигляді стопки тарілок, яка так і називається стопкою Гольджі або диктиосомою. Кожна диктиосома складається з 6-ти цистерн. Загальна кількість стопок Гольджі може бути різна – від 1 до 100. Завжди біля диктиосом групується маса дрібних одномембранних пухирців. Частина цих пухирців має більш складну будову. Їх мембрана вкрита додатковим шаром решітчастої оболонки. Ці пухирці отримали назву облямованих пухирців. Крім того розрізнюють більш крупні пухирці – секреторні пухирці, які ще називають гранулами або вакуолями. Вони містять речовини (секрети), що продукуються апаратом Гольджі (рис. 38).
Рис. 38. Тримірна схема реконструкції комплексу Гольджі тваринної клітини (зправа внизу – комплекс Гольджі під електронним мікроскопом.
Функції апарату Гольджі ще не зовсім вивчені. Вважається, що він є своєрідним “регулювальником” руху макромолекул в клітині. Більшість таких макромолекул, що синтезувалися на ЕПР, проходять через нього, набуваючи певної “зрілості”, тобто модифікуються. Це білки, глікопротеїни, гліколіпіди, компоненти клітинної стінки у рослин. Крім того білки, що призначені для секреції, пакуються в одномембранні секреторні пухирці, які потім зливаються з плазматичною мембраною. Звичайно такі пухирці багаторазового використання і здійснюють постійний кругообіг в клітині як “челноки”. Облямовані пухирці скоріш за все транспортують речовини до внутрішніх компартментів. Таким чином апарат Гольджі є другою ланкою після ЕПР в сортуванні, модифікації та транспорті синтезованих макромолекул (рис. 39).
Рис. 39. Схематичне зображення шляхів утворення вторинних лізосом різних типів. Від апарату Гольджі відокремлюються пухирці – первинні лізосоми, а потім в залежності від надходження до них ферментів та субстрату (інших речовин), вони утворюють або травні вакуолі при фагоцитозі, або мультівезикулярні тільця при ендоцитозі, або автофагуючі вакуолі. Слід зауважити, що точні шляхи таких перетворень невідомі.
Лізосоми та пероксисоми. Лізосоми були відкриті у 1949 році. Вони являють собою мембранний мішок, який заповнений ферментами. Відомо близько 40 таких ферментів. Це гідролітичні ферменти – протеази, нуклеази, ліпази, фосфатази, сульфатази тощо. Термін “гідролітичні” означає, що ферменти цього класу розщеплюють складні сполуки з одночасним приєднанням води. Цікаво, що всі вони діють в кислому середовищі. В мембрані лізосоми розташований транспортний білок, який “закачує” протони (Н+) і таким чином, створює кисле середовище з рН, що дорівнює 5-ти. У випадку руйнування лізосоми ферменти, що потрапляють до цитоплазми з рН значно вищою, стають неактивними. Так клітина захищає себе від можливих “аварій”. Деякі незаряджені малі молекули досить вільно проходять крізь мембрану лізосоми, а потім в кислому середовищі приєднують протон і стають зарядженими. Внаслідок цього молекула стає гідрофільною і не здатна тепер вільно вийти. Так відбувається їх нагромадження в лізосомі. Розрізнюють два основних класи лізосом. Це первинні лізосоми, які тільки відбрунькувалися від апарату Гольджі у вигляді облямованих пухирців і вторинні лізосоми – мембранні мішки, які містять набір ферментів та молекули інших речовин. Утворення вторинних лізосом відбувається шляхом злиття первинних. Такий процес багатоетапний і на різних його стадіях лізосомам дають різні назви. Наприклад, крупні травні вакуолі, ті, що утворюються при фагоцитозі крупних часток; так звані мультівезикулярні тільця, які містять багато пухирців; аутофагуючі вакуолі, які розщеплюють власні структури клітини (рис. 40).
Рис. 40. Утворення первинних та вторинних лізосом в клітині.
При деяких генетичних порушеннях ферментного складу лізосом спостерігається важкі порушення клітинного метаболізму. Прикладом може слугувати хвороба Хьорлера, при якій відбувається деформація кісток. У ферментному комплексі лізом відсутній фермент a - L –ідуронідаза, що викликає нагромадження в клітині цукристих речовин мукополіцукрів. Додавання цього ферменту приводить метаболізм до норми. Пероксисоми були виявлені лише на початку 1960-х років. Це також одномембранні мішечки, які утворюються на ЕПР. Часто пероксисоми залишаються з’єднаними з ЕПР тонким рукавом. Їх розміри близько 0,5 мкм. Як і мітохондрії цей компартмент засвоює кисень в клітині. Одна з гіпотез пояснює, що пероксисоми – це рудиментарний залишок дуже давнього органоїда, який потім був замінений мітохондріями. Але частина хімічних реакцій залишилася корисною для клітини і пероксисоми існують поряд з мітохондріями. Один з ферментів пероксисоми – каталаза – забезпечує важливу реакцію утилізації пероксиду водню (Н2О2), розщеплюючи його до води і кисню. Така реакція є рятівною для клітини, оскільки Н2О2 є сильним окисником. Вважають, що крупні пероксисоми печінки відіграють важливу роль у знешкодженні різних речовин. Наприклад, майже половина спожитого людиною етанолу окислюється тут до ацетальдегіду. Крім того, до половини всіх ліпідів клітини також розщеплюється в пероксисомах. Таким чином, одномембранні компартменти еукаріотичної клітини функціонально являють собою одне ціле. Так на ЕПР синтезуються білки та ліпіди, які сортируються, пакуються і потім у вигляді пухирців транспортуються до інших мембран. На ЕПР також утворюються пероксисоми. ЕПР тісно пов’язана з ядром та апаратом Гольджі. Саме в диктиосомах апарату Гольджі відбивається повторне сортування та пакування синтезованих продуктів. Апарат Гольджі також продукує лізосоми, які мають дуже високу ферментну активність і розкладають складні органічні речовини, що надходять як ззовні, так і власні відпрацьовані структури.
Поиск по сайту: |