|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Сукцесії в лісових біоценозах. ⇐ ПредыдущаяСтр 2 из 2
Дослідження сукцесій лісових біоценозів має практичне значення. Від їх осмислення та вміння прогнозувати залежить продуктивність лісів. У житті лісу фактор часу відіграє велику роль, зауважує П.С.Погребняк (1968), хоч і не очевидну для стороннього спостерігача. Темпи змін складу лісу найчастіше настільки повільні, що їх можна уявити лише складним шляхом різнобічних досліджень і за допомогою ряду абстракцій. При цьому найважливішою ознакою існування змін є зміна головних деревних порід. За масштабами часу в лісі спостерігаються принаймні три типи змін: 1. Зміна порід як наслідок онтогенезу деревостанів, тобто розвиток окремих поколінь лісу, починаючи із самосіву, підросту і закінчуючи старими, що досягли природної спілості, дерево станами [2]. 2. Зміна порід як наслідок стихійного втручання людини й інших зовнішніх факторів у житті лісу та процесу відновлення природного складу і структури лісу. Ці процеси триваліші, ніж згадані вище, оскільки вони охоплюють в найкращому випадку одне, два або три покоління деревостанів. 3. Зміна порід як наслідок крупномасштабних подій типу змін клімату, які відбуваються протягом тривалих відрізків часу. Концепцію зміни порід створив Г.Ф.Морозов, привівши в рух категорії, які здавались раніше незмінними, розкрив внутрішній звязок між лісами різного складу, показав, що одні з них є похідними від інших. Г.Ф.Морозов розкрив логіку сукцесій і зміну порід: смереки і сосни березою і осикою, дуба мяколистяними породами, сосни дубом, сосни і дуба смерекою, дуба супутніми породами (грабом). Як відомо, однією з основних причин зміни порід під час переходу корінних деревостанів у похідні виступає різниця в темпах росту у висоту. У випадку одночасного поселення і при сумісному рості цих порід панують швидкоростучі береза й осика, пригнічуючи дуб, смереку, бук і ялицю. Однак згодом корінні породи, що затримались у рості, "відвойовують" відведене їм природою місце і займають перший ярус біоценозу. Оселившись під наметом світлолюбних порід з їх характерною ажурною кроною, смерековий підріст з часом змикається зсередини (рис.3), що різко зменшує можливість відновлення недовговічних берези й осики. З часом, коли настає період їхньої природної стиглості (50-60 років), вони випадають, утворюючи ґрунт для смереки, яка запанує на всій території колишньої галявини. Однак коли смерекові крони піднімуться настільки, що почнуть проникати крізь листяний намет, "останній, пише П.С.Погребняк, найде нову "зброю" для боротьби з смерекою: листяні породи почнуть обхльостувати смерекові крони... І все ж, незважаючи на це, смерека продовжує рости і, нарешті, завдяки значній тривалості росту у висоту переганяє березу і осику" (1968). Смеречняки, відзначає П.С.Погребняк, господарі багатьох родючих місцезростань у лісовій зоні. Вони тут корінні насадження. Березняки й осичняки похідні насадження, які зявилися на місці корінних, як це було описано вище. Однак наявність їх не у всіх випадках свідчить про те, що територія раніше належала смереці. Березняки й осичняки можуть зявитися і на місці інших корінних типів насаджень сосняків, дібров, бучин, ялинників, модринників і т.д[15]. На основі конкурентних взаємодій видів у ході сукцесії відбувається поступове формування більш стійких комбінацій, відповідних конкретним абіотичних умов середовища. Приклад сукцесії, що приводить до зміни одного співтовариства іншим, – заростання невеликого озера з наступним появою на його місці болота, а потім лісу.(Див. додаток 2) Спочатку по краях озера утворюється сплавно – плаваючий килим з осок, мохів та інших рослин. Постійно озеро заповнюється відмерлими залишками рослин – торфом. Утворюється болото, яке поступово заростає лісом. Послідовний ряд поступово і закономірно змінюють один одного в сукцесії спільнот називається сукцесійною серією. Сукцесії в природі надзвичайно різномасштабні. Їх можна спостерігати в банках з культурами, що представляють собою планктонні співтовариства – різні види плаваючих водоростей і їх споживачів – коловерток, джгутикових в калюжах і ставках, на занедбаних ріллях, вивітрилися скелях і ін. В організації екосистем ієрархічність проявляється і в сукцессіонних процесах – більші перетворення біоценозів складаються з більш дрібних. У стабільних екосистемах з відрегульованим кругообігом речовин також постійно здійснюються локальні сукцесійні зміни, що підтримують складну внутрішню структуру спільнот.
Концепція клімаксу. Клімакс у екології та геоботаніці - заключне, відносно стійкий стан екосистем, що виникає в результаті змін, або сукцесій, і в значній мірі відповідне екологічним умовам певної місцевості. Клімакс залежить від кліматичних факторів, від місцевих особливостей грунтів і від впливів людини на природу. [1] Теоретично клімаксне співтовариство може підтримувати себе невизначено довго, всі внутрішні його компоненти врівноважені один з одним, і воно знаходиться в рівновазі з фізичним середовищем [2]. У польових умовах дуже складно виділити стійке клімаксовие співтовариство. Зазвичай вдається лише зауважити, що швидкість сукцесії падає до певного рівня, після якого спостерігач вже не помічає жодних змін. Період досягнення "клімаксової" стадії вимагає в різних спільнотах різного часу. Часто для завершення стабілізації необхідно 100-300 років, однак імовірність виникнення пожежі або урагану за цей час настільки висока, що сукцесія може ніколи не завершитись. Якщо не забувати, що лісові співтовариства північній помірної зони, а можливо, і тропіків все ще відновлюються після останнього льодовикового періоду, то постає питання, чи не є клімаксовою рослинність лише теоретичною [3]. У концепції клімаксу довга історія. Концепція клімаксу належить Фредеріку Клементсу (1916), який стверджував: в будь-якій кліматичній зоні існує лише один істинний клімакс — кінцевий стабільний стан рослинного угруповання, що перебуває в рівновазі з оточуючим середовищем [12]. До його виникнення ведуть усі сукцесії незалежно від того, почався він із піщаної дюни, перелогу чи навіть заростаючого ставка, що поступово переходить в суходільне угруповання. Тенслі (1939) та його однодумці заперечували Клементсу і говорили про полі-клімакс, стверджуючи при цьому, що клімакс на даній ділянці може визначатися одним або декількома факторами: кліматом, ґрунтовими умовами, топографією, пожежею тощо. Тому в одній кліматичній зоні, без сумніву, може існувати цілий ряд специфічних типів клімаксів (наприклад, букові праліси, чорновільшняки Закарпаття), склад яких є досить незмінним протягом тривалого часу (останніх двох століть). Поява клімаксових біоценозів пояснюється передусім зміною пануючих популяцій на основі конкурентної взаємодії, що приводить до поступового формування стійкіших угруповань, які відповідають існуючим абіотичним умовам середовища, які склалися за історичний період. Такий ланцюг біоценозів, які змінюють один одного, називають сукцесійним рядом, кожна тимчасова ланка якого являє собою певну стадію формування кінцевого клімаксового угруповання (Див.додаток. 4). Сукцесія рослинності - це послідовний ряд зміни серійних (тимчасово існуючих) рослинних співтовариств на конкретному місцеперебуванні після виведення конкретної экосистеми зі стану динамічної рівноваги. У результаті сукцесії на конкретному місцеперебуванні відновлюється вихідне рослинне співтовариство, назване геоботаніками клімаксовим, або корінним. Корінне співтовариство рослин стійко й у даних кліматичних умовах не змінюється. При цьому конкретна екосистема повертається у свій вихідний стан і перебуває в ньому доти, поки не зміняться клімат, рельєф, гідрологічний режим, поки знову не пройде пожежа, або не случиться якась інша катастрофа. І знову почнеться нова сукцесія, що або приведе до відновлення вихідного співтовариства, або немає. Якщо пройшов пожежу, ліс вирубали, луг або степ розорали і потім закидали, то, швидше за все, сукцесія завершиться відновленням корінного співтовариства, що так само, як і вихідне, буде знаходитися в стані відносної рівноваги з зовнішнім середовищем. Однак, якщо змінився клімат, понизився або підвищився рівень ґрунтових вод, співтовариство рослин, виведене зі стану рівноваги, у процесі сукцесії не відновиться колишнім. Воно буде корінним, согласуясь з новими умовами середовища, але іншим, чим вихідне корінне співтовариство [9]. Існує гіпотеза, припущення, що в сучасних умовах у районах з хитливим мусонним кліматом корінні співтовариства взагалі не встигають сформуватися. Істотні для рослинності переміщення клімату відбуваються швидше, ніж триває сукцесійний ряд. Висновки 1. Поступові необоротні зміни складу та структури біогеоценозу, що спричинюються зовнішніми або внутрішніми факторами, називають сукцесіями. Класичний приклад сукцесії — заростання озера й перетворення його на болото[13]. 2. Сукцесії бувають повільними (тривають тисячоліття й десятки тисяч років), середніми (століття) й швидкими (десятиліття). Вони можуть відбуватися з внутрішніх причин (наприклад, утворення нових видів) або під впливом зовнішніх факторів (наприклад, засолення, підтоплення, вселення видів «чужих» фаун та флор), мати природне (наприклад, у результаті підняття чи опускання суші) чи антропогенне (вирубування лісу, розорювання степу) походження, бути прогресуючими (тобто супроводжуватися збільшенням продуктивності й видового багатства, як у випадку заростання новоутворених річкових піщаних кіс) або, навпаки, регресуючими. В.М.Сукачов виділив чотири види сукцесій: 1) сингенетичні; 2) ендоекогенетичні; 3) екзогенетичні; 4) гологенетичні. 3. Первинна сукцесія - це заростання місця, раніше не зайнятого рослинністю: голих скель або застиглої вулканічної лави. Вторинна сукцесія формується в результаті втручання діяльності людини в природні процеси. 4. Сукцесійні зміни звичайно зв'язують з тим, що існуюча экосистема (співтовариство) створює несприятливі умови для її організмів, що наповняють, (ґрунтовтому, неповний кругообіг речовин, самоотруєння продуктами виділення або розкладання і т.п.). Такі явища реальні, але не пояснюють усіх випадків змін екосистем. Поряд із природними факторами причинами динаміки екосистем усі частіше виступає людина. До дійсного часу їм зруйнована більшість корінних (клімаксних) екосистем. Наприклад, степи майже цілком розорані (збереглися тільки на заповідних ділянках). Переважні площі лісів представлені перехідними (тимчасовими) екосистемами з листяних деревних порід (береза, осика, рідше верба, вільха й інші). Ці ліси звичайно називають похідними, або вторинними. Для будь-якої сукцесії, особливо первинної, характерні наступні загальні закономірності протікання процесу: 1. На початкових стадіях видова розмаїтість незначна, продуктивність і біомаса малі. але в міру розвитку сукцесії ці показники зростають. 2. З розвитком сукцесійоного ряду збільшуються взаємозв'язки між організмами. Особливо зростає кількість і роль симбіотичних відносин. Повніше освоюється середовище існування, ускладнюються ланцюги і мережі харчування[14]. 3. Зменшується кількість вільних екологічних ніш, і в клімаксному співтоваристві вони або відсутні, або знаходяться в мінімумі. У зв'язку з цим у міру розвитку сукцесій зменшується імовірність спалахів чисельності окремих видів. 4. Інтенсифікуються процеси кругообігу речовин, потік енергії екосистем. 5. Швидкість суцесійоного процесу в більшій мері залежить від тривалості життя організмів, що відіграють основну роль у додаванні і функціонуванні екосистем. У цьому відношенні найбільш тривалі сукцесії в лісових екосистемах. Коротші вони в екосистемах, де автотрофна ланка представлена трав'янистими рослинами, і ще швидше протікають у водних екосистемах. 6. Незмінюваність завершальних (клімаксних) стадій сукцесій відносна. Динамічні процеси при цьому не припиняються, а лише сповільнюються. Продовжуються динамічні процеси, зумовлювані змінами сереої питомої ваги займають динамічні процеси циклічного (флуктуаційного) плану. 7. У зрілій стадії клімаксного співтовариства біомаса звичайно досягає максимальних або близьких до максимальних значень. Неоднозначна продуктивність окремих співтовариств на стадії клімаксу. Звичайно вважається, що в міру розвитку сукцесійонного процесу продуктивність збільшується і досягає максимуму на проміжних стадіях, а потім у климаксном співтоваристві різко зменшується. Останнє зв'язують, по-перше, з тим, що в цей час максимум первинної продукції споживається консументами, а, по-друге, екосистема розвиває надзвичайно велику масу асиміляційного апарату, що веде до дефіциту освітленості, наслідком чого є зниження інтенсивності фотосинтезу при одночасному зростанні втрат продуктів асиміляції на дихання самих автотрофів [9]. Ці положення не можна поширювати на всі клімаксні співтовариства. Наприклад, немає реальних передумов для збільшення чисельності гетеротрофів у хвойних лісах у порівнянні з листяними. Скоріше, в останніх більше споживачів зеленої продукції і, імовірніше, спалаху чисельності окремих виглядів-фітофагів, наприклад, комах. Немає також ні теоретичних передумов, ні фактичних даних, які б свідчили, що в зрілої климаксної системи, наприклад у ялинових лісах, маса хвої досягає надмірно високих значень. Це суперечить принципам адаптації до збільшення біогенної геохімічної енергії організмами як умові їх виживання (другий біогеохімічний принцип В.І. Вернадского). Весь досвід лісівництва також свідчить про найбільш високу продуктивність клімаксних лісових співтовариств (стосовно до лісової зони хвойних або змішаних хвойно-листяних лісів). У противному випадку, з погляду одержання продукції (деревини), неминучий висновок про недоцільність орієнтації на вирощування і збереження клімаксних стадій лісів. Стосовно до іншим екосистем, наприклад лучних, можна погодитися з тим, що можливості одержання продукції на клімаксній стадії зменшуються, однак не тому, що скорочується її наростання (приріст, продуктивність), а з тієї причини, що більш значна частина її відчужується гетеротрофами в результаті утворення стійких ланцюгів виїдання. Іншими словами, продуктивність екосистем на клімаксних стадіях сукцесій висока. Як правило, максимальна внаслідок більш повного освоєння простору. Однак можливості зняття людиною первинної продукції лімітуються (іноді до нульових значень) унаслідок включення її в ланцюзі харчування[16].
Література 1. Білявський Г. О., Падун М. М., Фурдуй Р. С. Основи загальної екології. — К.: Либідь. 1995 — 368 с. 2. Білявський Г. О., Фурдуй Р. С. Практикум із загальної екології. // Навч. посібн. — К.: Либідь, 1997. — 160с. 3. Волошин І. М. Методика дослідження проблем природокористування. — Львів: ЛДУ, 1994. — 160 с. 4. Гайнріх Д., Гергт М. Екологія. - К.: Знання-прес, 2001. – 287 с. 5. Голубець М.А., Кучерявий В.П., Генсірук С.А. та ін. Конспект лекцій з курсу "Екологія і охорона природи". - К., 1990. 6. Екологічний словник: Навч. посібник /В.В.Прежко та ін. – Харків: ХДАМГ, 1999. – 416 с. 7. Екологія і закон: Екологічне законодавство України. У 2-х кн./ Відповідальний редактор док. юрид. наук, професор, акад. Андрейцев В. А. — К.: Юрінком їнтер, 1997. — 704 с. 8. Заверуха Н.М., Серебряков В.В., Скиба Ю.А. Основи екології. – К.: Каравела, 2006. - 368 с. 9. Запольський А.К., Салюк А.І. Основи екології. – К.: Вища школа, 2003. – 358 с. 10. Злобін Ю.А. Основи екології.- К.: Лібра, 1998. – 249. 11. Корсак К.В., Плахотнік О.В. Основи екології. - К.: МАУП, 2000. – 238 с. 12. Кучерявий В.П. Екологія, - Львів: Світ, - 500 с. 13. Одум Ю. Экология. В 2 т. - М., 1986., Т 1. – 328 с. 14. Одум Ю. Экология. В 2 т. - М., 1986., Т.2. – 376 с. 15. Основи екології та соціоеколотії. // Навч. посібн. під. ред. Назарука М.М. — Львів: "За вільну Україну". - 1997. — 210 с. 16.Інтернет джерела׃ uk.wikipedia.org eduknigi.com ecotime.info bibliofond.ru
Додатки Додаток 1
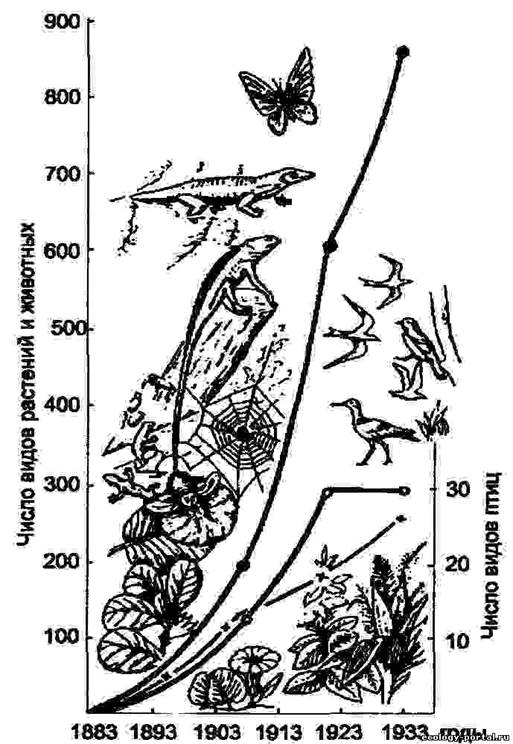
Динаміка видового різноманіття на о. Кракатау після повного знищення аборигенної флори і фауни вулканічним вибухом в 1893 р. (за Р. Макартур та Е. О. Вильсону, 1967)
Додаток 2
Сукцесія при заростанні невеликого озера (За А.О. Рувінська та ін, 1993)
Додаток 3
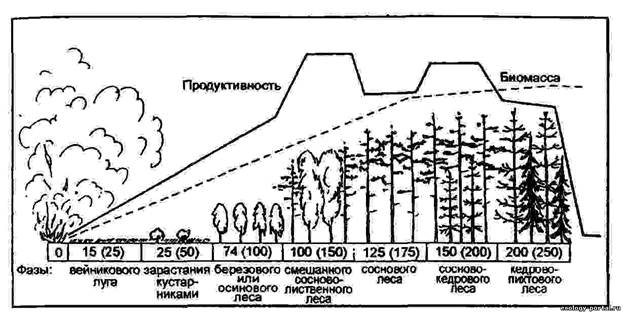
Вторинна сукцесія сибірського темно-хвойного лісу (Ялицево-кедрової тайги) після спустошливого лісової пожежі (За М. Ф. Реймерс, 1990) Примітка: числа в прямокутниках – коливання в тривалості проходження фаз вторинної сукцесії (в дужках вказано термін їх закінчення). Біомаса та біологічна продуктивність показані в довільному масштабі (криві відображають якісну і кількісну сторони процесу). Додаток 4
Зміна видового різноманіття і чистої первинної продукції в ході сукцесії, що веде до встановлення клімаксового мішаного лісу із сосни та дуба: 1 — трав'яні рослини, 2 — чагарники, 3 — сосна, 4 — дуб.
Поиск по сайту: |