|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
IV. Хроматин, его классификация. Строение хромосом ⇐ ПредыдущаяСтр 2 из 2
В ядре клеток обнаруживаются мелкие зерна и глыбки материала, который окрашивается основными красителями и поэтому был назван хроматином (от греч. chroma – краска). Хроматин – это деспирализованная форма существования хромосом в неделящемся ядре. Его химическую основу составляет дезоксирибонуклеопротеин – комплекс ДНК с гистоновыми и негистоновыми белками. При этом до момента репликации ДНК каждая хромосома содержит лишь одну линейную молекулу ДНК. Хроматин соответствует хромосомам, которые в интерфазном ядре представлены длинными перекрученными нитями и неразличимы как индивидуальные структуры. Выраженность спирализации каждой из хромосом неодинакова по их длине. Реализацию генетической информации осуществляют деспирализованные участки хромосом. Белки составляют значительную часть вещества хромосом. На их долю приходится около 65 % массы этих структур. Все хромосомные белки разделяются на две группы: гистоны и негистоновые белки. Гистоновые белки включают 5 главных видов белков: Н1, Н2А, Н2В, Н3 и Н4 (Н – от histon). Гистоны первых трех классов (Н1, Н2А, Н2В) содержат большое количество аминокислоты лизина. В состав гистонов Н3 и Н4 входит много аминокислоты аргинина. Гистоны - это положительно заряженные основные белки, которые достаточно прочно соединяются с молекулами ДНК, фосфатные группы которых несут отрицательный заряд. Связь гистонов с ДНК препятствуют считыванию заключенной в ДНК биологической информации. В этом состоит их регуляторная роль. Кроме того, эти белки выполняют структурную функцию, обеспечивая пространственную организацию ДНК в хромосомах. Общее массовое содержание кислых (негистоновых) белков в хромосомах существенно меньше, чем гистонов. Однако эти белки чрезвычайно разнообразны (число фракций негистоновых белков превышает 100). Вероятно, некоторые из кислых белков играют структурную роль, участвуя в образовании наднуклеосомных уровней укладки хромосом. Другую группу составляют многочисленные ферменты, обеспечивающие процессы репликации, модификации, репарации и транскрипции. Самой разнообразной по составу, видимо, является группа регуляторных белков. Они контролируют активность вышеуказанных ферментов, а также доступность тех или иных участков ДНК для этих ферментов.
Классификация хроматина. В ядрах абсолютного большинства клеток генетический материал представлен диффузно расположенным хроматином. Тем не менее, при окраске хроматина уже под световым микроскопом обнаруживается его неоднородность. Основная масса хроматина, имеющая бледную окраску, получила название эухроматина. Кроме эухроматина, в составе хроматина ядра выявляются участки хроматина с более темной окраской. Такой вариант хроматина называют гетерохроматином. (Эухроматин и гетерохроматин отличаются друг от друга по степени спирализации. Гетерохроматин конденсирован более сильно, поэтому и окрашивается более интенсивнее эухроматина.) Итак, различают два вида хроматина: 1) эухроматин (от греч. eu – хорошо, полностью и хроматин), участки хромосом, сохраняющие деспирализованное состояние в покоящемся ядре (в интерфазе) и спирализующиеся при делении клеток (в профазе); эухроматин соответствует сегментам хромосом, локализующихся ближе к центру ядра. Эухроматин больше деспирализованный, менее компактный, содержит большинство генов и потенциально способен к транскрипции. Предполагается, что в нем сосредоточена та ДНК, которая в интерфазе генетически активна. Эухроматин отличается от гетерохроматина меньшим содержанием метилированных оснований и блоков повторяющихся последовательностей ДНК, большим количеством негистоновых белков и ацетилированных молекул гистонов, менее плотной упаковкой хромосомного материала, что, как полагают, особенно важно для активности эухроматина и делает его потенциально более доступным для ферментов, обеспечивающих транскрипцию. Эухроматин может приобретать свойства факультативного гетерохроматина – инактивироваться, что является одним из способом регуляции генной активности. 2) гетерохроматин часть хроматина, находящаяся в плотно спирализованном, упакованном состоянии в течение всего клеточного цикла. Гетерохроматин соответствует конденсированным, плотно скрученным сегментам хромосом (что делает их недоступными для транскрипции). Он интенсивно окрашивается основными красителями и в световом микроскопе имеет вид темных пятен, гранул. Гетерохроматин располагается ближе к кариолемме, более компактен, чем эухроматин и содержит «молчащие» гены, т.е. гены, которые в настоящий момент неактивны. Гетерохроматичные районы хромосом, как правило, реплицируются позже эухроматиновых и не транскрибируются, т.е. генетически весьма инертны. Ядра активных тканей и эмбриональных клеток большей частью бывают бедны гетерохроматином. Различают конститутивный и факультативный гетерохроматин. 2.1) конститутивный (структурный) хроматин впервые был обнаружен в начале 30-х годов Гейтцем, который заметил, что в интерфазных ядрах существуют постоянные участки конденсированного хроматина. Его наличие не зависит от степени дифференцированности ткани или от функциональной активности. Гетерохроматин – это компактные участки хромосом, которые в профазе появляются раньше других частей в составе митотических хромосом и в телофазе не декондесируются, переходя в интерфазное ядро в виде интенсивно красящихся плотных структур (хромоцентров), которые располагаются неподалеку от кариолеммы. Постоянно конденсированными зонами чаще всего являются центромерные и теломерные участки хромосом. Конститутативный хроматин не транскрибируется, реплицируется позже всего остального хроматина, в его состав входит (сателлитная) ДНК, обогащенная часто повторяющимися последовательностями нуклеотидов; служит для взаимодействия хромосом с ламиной.
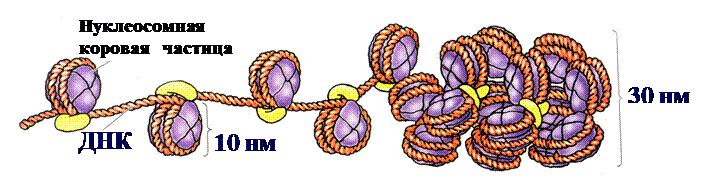
2.2) факультативный (функциональный) гетерохроматин обнаруживается при сравнении ядер разных клеток одного организма, при этом выявляется, что определенные участки хроматина в одних клетках могут быть гетерохроматиновыми, а в других эухроматиновыми. В ДНК факультативного гетерохроматина локализованы гены, которые неактивны из-за его сильной конденсации. Тем не менее, эти гены способны функционировать, если данный район хроматина переходит в деконденсированное (эухроматиновое) состояние. Таким образом, факультативный гетерохроматин представляет собой отражение одного из способов регуляции действия генов – с его помощью в различных клетках можно «выключать» разные гены. Кроме того, факультативный гетерохроматин может присутствовать только в одной из гомологичных хромосом. Пример гетерохроматина такого типа – вторая Х-хромосома у женских особей млекопитающих, которая в ходе раннего эмбриогенеза инактивируется вследствие ее необратимой конденсации. Так, у человека сначала функционируют две Х-хромосомы (у ♀♀), что необходимо для нормального протекания оогенеза (развития женских половых клеток), на 16-е сутки во всех клетках женского эмбриона одна из Х-хромосом образует тельце полового хроматина (тельце Бара), которое может быть обнаружено вблизи ядерной мембраны интерфазных клеток в виде хорошо окрашивающегося гетерохроматинового образования. Уровни компактизации хроматина. Сохраняя преемственность в ряду клеточных поколений, хромосомы в зависимости от периода и фазы клеточного цикла меняют свое строение. В интерфазе они образуют хроматин. При переходе клетки к митозу, особенно в метафазе, хроматин приобретает вид хорошо различимых отдельных интенсивно окрашенных телец – хромосом. Интерфазную и метафазную формы существования хромосом расценивают как два полярных варианта их структурной организации, связанных в клеточном цикле взаимопереходами. Различают следующие уровни компактизации ДНК: 0) Двойная спираль ДНК представлена «голой» ДНК, не связанной с белками. Ширина двойной спирали ДНК составляет 2 нм. 1) Нуклеосомный уровень хроматина возникает при взаимодействии молекулы ДНК с молекулами белков-гистонов. Два с половиной витка двойной спирали ДНК (в146-200 пар нуклеотидов) наматываются снаружи на белковый кор, образуя нуклеосому (рис. 9,10).
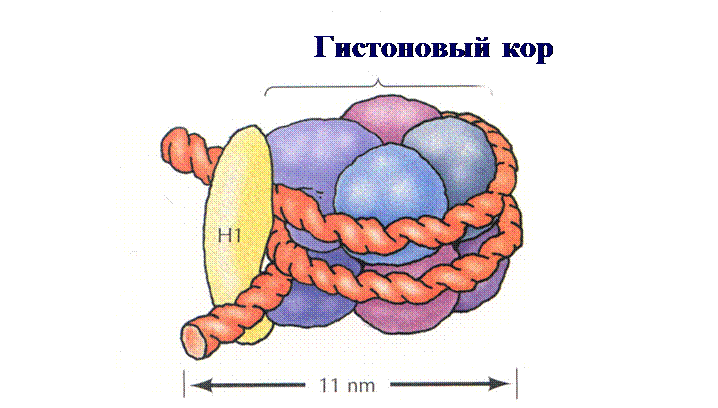
Кор – это белковый октамер, состоящий из 8-ми гистоновых белков четырех типов (Н2А, Н2В, Н3, Н4). Каждый гистон представлен двумя молекулами. ДНК наматывается на кор снаружи, образуя два споловиной витка (рис. 10). Участок ДНК между нуклеосомами называется линкером и имеет протяженность 50-60 пар нуклеотидов. Толщина нуклеосомной фибриллы (нити)составляет 8-11 нм.
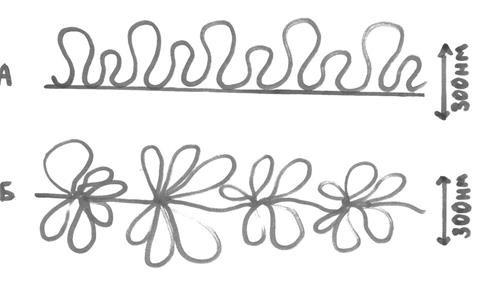
Рис. 10. Структура нуклеосомной коровой частицы. 2) Нуклеомерный (хроматиновая фибрилла, или нить). Нуклеосомная структура закручивается, образуя суперспираль. В ее образовании принимает еще один гистоновый белок Н1, лежащий между нуклеосомами и связанный с линкером. К каждому линкеру присоединяется 1 молекула гистона Н1. Молекулы Н1 в комплексе с линкерами взаимодействуют между собой и вызывают суперспирализацию нуклеосомной фибриллы. В результате образуется хроматиновая фибрилла (рис. 11), толщина которой составляет 30 нм:
Рис. 11. Хроматиновая фибрилла. На нуклеомерном уровне ДНК компактизована в 40 раз. Суперспирализация происходит двумя способами. Нуклеосомная фибрилла может образовывать спираль второго порядка, которая имеет форму соленоида. При втором варианте суперспирализация 8-10 нуклеосом образуют крупную компактную структуру – нуклеомеру. В обоих случаях формируется новый уровень пространственной организации хроматина, который называют нуклеомерным уровнем. Этот уровень не допускает синтеза РНК с нуклеомерной ДНК (на нуклеомерном уровне организации хроматина транскрипция не происходит).
Рис. 12 Петельная структура хроматина. 4) Хромонемный(от chroma – краска, nema – нить) уровень. Хроматин является субстанцией, которая образует хромосомы. В простейшем случае хромосома содержит одну целостную гигантскую молекулу ДНК в комплексе с белками, т.е. фибриллу ДНП. Такая ДНП-фибрилла называется хромонемой. Хромонемный уровень образуется в результате сближения хромомеров по длине. Перед делением клетки, в S-период интерфазы, каждая хромосома, содержащая одну хромонему, удваивается и состоит из двух хромонем. Эти хромонемы соединены в определенном участке хромосомы специальной структурой – центромерой.
5) Хроматидный уровень формируется в результате укладывания хромонемы в очень компактную структуру – хроматиду, при этом хромонема образует множество дополнительных петель. Таким образом, хромонема складывается несколько раз, образуя тело хроматиды. Толщина хроматиды около 700 нм. В норме каждая хроматида содержит одну хромонему, но очень плотно упакованную. Хроматиду можно назвать нереплицированной хромосомой. После репликации ДНК хромосома содержит 2 хроматиды.
Метафазная хромосома состоит из двух хроматид (рис. 15 Е). Толщина ее составляет 1400 нм. Хроматиды соединены центромерой. При делении клетки хроматиды расходятся и попадают в разные дочерние клетки. Последовательность компактизации хроматина, начиная с молекулы ДНК до хромосомы можно проследить на рисунке 15.
Рис. 15. Уровни компактизации хроматина:
Эухроматину соответствует нуклеосомный и нуклеомерный уровни компактизации ДНК. Гетерохроматину – хромомерный и хромонемный уровни компактизации ДНК, а хроматидный и хромосомный уровни наблюдаются во время митоза. Таким образом, хроматин и хромосомы представляют собой дезоксирибонуклеопротеиды (ДНП), но хроматин* – это раскрученное, а хроматиды, следовательно, и хромосомы – скрученное состояние. Хроматид и хромосом в интерфазном ядре нет, они появляются при разрушении ядерной оболочки (во время деления: на стадиях поздней профазы, метафазы, анафазы, ранней телофазы). * Термин хроматин употребляется также для обозначения наследственного вещества клетки, представляющего собой дезоксирибонуклеопротеидный комплекс различной степени компактизации. Строение хромосом Хромосомы представляют собой наиболее упакованное состояние хроматина. Наиболее компактные хромосомы видны на стадии метафазы, при этом они состоят из двух хроматид, связанных в области центромеры. Хроматиды генетически идентичны, они образуются во время репликации и поэтому называются сестринскими хроматидами.

Рис. 16. Метафазная хромосома.
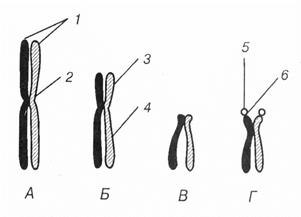
Рис. 17. Типы хромосом.
V. Ядрышко Ядрышко – это хорошо заметная в световой микроскоп округлая структура, является самой плотной структурой ядра. Ядрышко находится внутри ядра. Ядрышко интенсивно окрашивается ядерными красителями, т.к. содержит большое количество РНК и ДНК. В состав ядрышка входят рибонуклеопротеиды (РНП). В ядрах клеток эукариот может быть одно, два или несколько ядрышек. Ядрышко – это не отдельная от хроматина структура, а его производная. Ядрышко лишено мембраны и образуется вокруг участков хромосом, в ДНК которых закодирована информация структуре р-РНК. Эти специализированные структуры (петли) хромосом носят название ядрышковых организаторов. Ядрышковые организаторы расположены в области вторичной перетяжки спутничных хромосом. На ДНК ядрышкового организатора синтезируются р-РНК. Обычно функцию ядрышкового организатора выполняет вторичная перетяжка спутничных хромосом. У человека такие участки имеются в 5-ти хромосомах – 13-й, 14-й, 15-й, 21-й и 22-й, где располагаются многочисленные копии генов, кодирующих рибосомальные РНК (р-РНК). Размеры и число ядрышек увеличивается при повышении функциональной активности клетки. Особенно крупные ядрышки характерны для эмбриональных и активно синтезирующих белки клеток, а также для клеток быстрорастущих злокачественных опухолей. Ядрышко исчезает в профазе митоза, когда ядрышковые организаторы «растаскиваются» в ходе конденсации соответствующих хромосом, вновь формируясь в телофазе. Функции ядрышка заключаются в синтезе р-РНК и ее сборке в предшественники рибосомальных субъединиц. Под электронным микроскопом в ядрышке обнаруживают две области: 1) фибриллярная область состоит из множества тонких нитей (5-8 нм) и располагаются во внутренней части ядрышка. Здесь же располагаются участки ДНК ядрышковых организаторов. В фибриллярной части ядрышка происходит образование р-РНК в процессе транскрипции, созревание (процессинг) р-РНК. 2) глобулярная часть (гранулярный компонент) образована скоплением плотных частиц диаметром 10-20 нм. В глобулярной части происходит объединение р-РНК с белками, поступившими из цитоплазмы, т.е. происходит образование субъединиц рибосом. Фибриллярный и гранулярный и гранулярный компоненты ядрышка образуют т.н. ядрышковую нить (нуклеосому) толщиной 60-80 нм, которая в пределах ядрышка формирует широкопелистую сеть, выделяющуюся большей плотностью на фоне менее плотного матрикса. Ядрышко окружено перинуклеальным хроматином, небольшое количество хроматина проникает с периферии внутрь ядрышка (интрануклеолярный хроматин). В ядрышке клеток обнаруживаются мелкие зернышки и глыбки хроматина, который окрашивается основными красителями; состоит из комплекса ДНК и белка и соответствует хромосомам, которые в интерфазном ядре представлены длинными тонкими перекрученными нитями и неразличимы как визуальные структуры.
Поиск по сайту: |




 А - нуклесомная фибрилла, Б - элементарная хроматиновая фибрилла; В - интерфазная петельная структура, Г- хромонема; Д – хроматида; Е - метафазная хромосома.
А - нуклесомная фибрилла, Б - элементарная хроматиновая фибрилла; В - интерфазная петельная структура, Г- хромонема; Д – хроматида; Е - метафазная хромосома.