|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
ФУНКЦИОНАЛЬНЫЕ СВОЙСТВА БЕЛКОВ
Растительные белки находят применение в производстве пищевых продуктов в качестве ингредиентов питательной, технологической и лечебно-профилактической значимости благодаря присущим им уникальным функциональным свойствам. Понятие "функциональные свойства белков" впервые ввели Серкл и Джонсон в 1962 г. Под функциональными свойствами понимают физико-химические характеристики белков, определяющие их поведение при переработке в пищевые продукты и обеспечивающие определенную структуру, технологические и потребительские свойства (В. Толстогузов, 1987). К наиболее важным функциональным свойствам белков относятся растворимость, водосвязывающая и жиросвязывающая способность, способность стабилизировать дисперсные системы (эмульсии, пены, суспензии), образовывать гели, пленкообразующая способность, адгезионные и реологические свойства (вязкость, эластичность), способность к прядению и текстурированию. Белки с высокими функциональными свойствами хорошо растворяются в воде, образуют прочные гели, стабильные эмульсии и пены; белки с низкими функциональными свойствами не набухают в воде, не способны образовывать вязкие, эластичные массы, гели, не стабилизируют пены и эмульсии. Некоторые известные белки не попадают под указанные выше закономерности. Так, белки пшеничной клейковины, несмотря на низкую растворимость в воде (2–5%), образуют структурные коллоидные системы – гели, которые выдерживают нагревание, замораживание и сушку, а белки из отрубей и тритикале с растворимостью 10–20% обладают высокими жироэмульгирующими и пенообразующими свойствами. Отклонения от указанных выше закономерностей объясняются трудностью создания в экспериментах при оценке функциональных свойств стандартных условий, так как белки имеют разные оптимальные значения свойств, а модельные системы не учитывают многофункциональность и взаимодействие белков с другими компонентами пищи (липидами, углеводами и т. д.). Поэтому признано, что функциональные свойства белков должны выражаться не только в численных значениях, но и в профилях зависимостей от технологических или других факторов. Данный подход в оценке функциональных свойств нашел отражение в применении для белков нового термина – "технофункциональные", включающего особенности технологических процессов при производстве, хранении и потреблении пищевых продуктов. При этом функциональные свойства белков оцениваются для конкретных пищевых систем в рамках выбранного направления путем сравнения их со свойствами традиционных или других известных белков. Растворимость, являясь первичным показателем оценки функциональных свойств белков, характеризуется коэффициентами КРА и КДБ. В первом случае определяют количество азота, во втором – количество белка, перешедшего в раствор (в процентах от общего содержания его в продукте). Специфическая последовательность аминокислотных остатков в полипептидных цепях, неравномерное расположение гидрофобных и гидрофильных групп на поверхности белков, наличие или отсутствие спирализованных участков обуславливают особенности функциональных свойств. Растворимость в наибольшей степени зависит от присутствия нековалентных взаимодействий: гидрофобных, электростатических и водородных связей (см. Строение пептидов и белков). При рассмотрении гидрофобных взаимодействий белков различают среднюю и относительную (поверхностную) гидрофобность. Под средней гидрофобностью понимают энергию стабилизации, приходящуюся на одну неполярную боковую группу при связывании ее внутри глобулы белковой молекулы, под относительной – степень гидрофобного взаимодействия неполярных остатков аминокислот, расположенных на поверхности глобул. Последний вид гидрофобности оценивается по связыванию с липидами и углеводами, распределению белков в двухфазных водных системах, содержащих полимеры с разной гидрофобностью, и т.д. За счет относительной гидрофобности осуществляется взаимодействие с липидами и формируется четвертичная структура белков. Чем ниже относительная гидрофобность белков (то есть ниже взаимодействие между глобулами и выше сила отталкивания), тем выше взаимодействие их с молекулами растворителя, следовательно выше растворимость. Вклад электростатических сил в растворимость белков зависит от рН среды и присутствия солей. При рН, соответствующем ИЭТ, белки имеют наименьшую растворимость, так как суммарный заряд на их молекулах равен нулю и частицы лишены способности отталкиваться за счет электростатических взаимодействий с молекулами растворителя. В кислой или щелочной среде, наоборот, обеспечивается взаимодействие противоположно заряженных ионов растворителя (Н+ или ОН- соответственно) с поверхностями белковых частиц, заряженных положительно в кислой среде и отрицательно в щелочной, а значит и переход белков в раствор. В кислой среде белок имеет положительный заряд вследствие подавления диссоциации карбоксильных (–СООН) групп, в щелочной – отрицательный за счет подавления диссоциации основных (–NH2) групп. Зависимость растворимости белков от концентрации солей носит нелинейный характер. При добавлении небольших количеств солей растворимость увеличивается, так как ионы препятствуют электростатическому взаимодействию боковых групп белка между собой. Высокие концентрации солей, снижающие гидратацию полипептидных цепей, наоборот, усиливают гидрофобные белок-белковые взаимодействия и вызывают выпадение белка в осадок (высаливание). Использование в качестве растворителя воды, разбавленных растворов солей, щелочей и водно-спиртовых растворов обеспечивает перевод гетерогенных смесей в раствор, соответственно, альбуминов, глобулинов, глютелинов и проламинов и получение белковых фракций, различающихся по аминокислотному составу, молекулярным массам и функциональным свойствам. Различия в растворимости белков пищевого сырья лежат в основе технологических процессов выделения изолятов и концентратов и имеют непосредственное отношение к качеству многих пищевых продуктов. Важное значение растворимость белков имеет для повышения качества пищевых продуктов, в производстве которых предусмотрен их гидролиз (автолиз) и денатурация (начальные технологические стадии, сушка и хранение). Потеря растворимости, как правило, сопровождается изменением и других важных функциональных свойств, что в значительной мере отражается на качестве продуктов и степени перевариваемости белка в желудочно-кишечном тракте. Особые требования к растворимости белков предъявляются при использовании последних в производстве напитков, хлебных, мучных кондитерских и макаронных изделий. В напитках применяются белки с высокой растворимостью, в изделиях из муки – с низкой. Применение белков с чрезмерно высокой растворимостью в составе хлебопекарных улучшителей отрицательно отражается на эластично-вязкоупругих свойствах теста. Незначительное количество растворимого белка должно содержаться в текстурированных формах белка, зерновых продуктах, приготовленных высокотемпературной экструзией, и макаронных изделиях. Свойства белковых суспензий. При использовании белков в качестве обогатителей, наполнителей (разбавителей), функциональных ингредиентов и аналогов мясных и рыбных изделий большое значение имеют такие свойства белковых суспензий, как ограниченная степень набухания и размер частиц, водо- и жиросвязывающая способность, адгезионные свойства, значение рН и буферная емкость, образование вязко-упругоэластичных масс и гелей. Водосвязывающая способность характеризуется адсорбцией воды при участии гидрофильных остатков аминокислот, жиросвязывающая – адсорбцией жира за счет гидрофобных остатков. При невысокой влажности гидрофильные группы, взаимодействуя с молекулами воды, образуют мономолекулярный слой, при высокой – вокруг глобул белка формируется многослойная структура с одновременным проникновением воды во впадины и выступы. Общее количество воды и жира на поверхности достигает 0,2–0,4 г на 1 г белков. Способность белков удерживать жир и воду зависит не только от особенностей аминокислотного состава и структуры, но и от фракционного состава, способа обработки, рН среды, температуры и присутствия углеводов, липидов и других белков. В пшеничном тесте при добавлении соевого белка или пшеничной клейковины водопоглотительная способность положительно коррелирует с количеством нерастворимой фракции белков и отрицательно – с содержанием растворимой. Высокая способность белков удерживать воду в пищевых продуктах (мясных, хлебобулочных и т.д.) повышает выход последних, удлиняет сроки хранения и улучшает текстуру. Денатурированные белки имеют пониженную водосвязывающую способность, и их применение отрицательно сказывается на качестве хлеба. Высокая жироудерживающая способность белков обеспечивает нежную и однородную текстуру изделий, исключает отделение жира, сморщивание изделий, уменьшает потери при варке и жарении. Жироэмульгирующая и пенообразующая способности белков широко используются в практике получения жировых эмульсий и пен. Присутствие в одной белковой цепи гидрофобных и гидрофильных группировок обеспечивает распределение молекул определенным образом на границе раздела фаз вода–масло и вода–газ. Ориентация гидрофильных групп белка к воде, а гидрофобных – к маслу на границе раздела фаз в виде прочного адсорбционного слоя снижает поверхностное натяжение в дисперсных системах и делает их агрегативно устойчивыми и одновременно вязкими. Наиболее широко распространены пищевые эмульсии "масло в воде" (м/в) и "вода в масле" (в/м), называемые, соответственно, прямыми и обратными. В производстве новых форм белковой пищи большое значение приобрели и эмульсии "вода в воде" (в/в). Все виды эмульсий с белком получают механическим диспергированием одной жидкости в другой при помощи мешалок, гомогенизаторов, обеспечивающих в поле сил сдвига деформацию дисперсионной среды с образованием мелких частиц. Эмульгирующие свойства белков оценивают по эмульгирующей способности, эмульгирующей емкости, стабильности эмульсий и т.д., описание которых можно найти в специальной литературе. Пены (дисперсные системы с газообразной фазой и жидкой или твердой средой) получают механическим распределением воздуха в растворе белка путем взбивания или за счет вскипания воды, понижения давления, обеспечения химических и микробиологических процессов в белоксодержащих пищевых системах. Так, белки клейковины образуют пену в хлебном тесте под действием диоксида углерода при брожении, а в кондитерском – за счет химических разрыхлителей при выделении аммиака и диоксида углерода. Пенообразующие свойства белков характеризуются пенообразующей способностью и стабильностью пены. Первый показатель измеряется объемом пены, отнесенным к массе белка, второй – периодом ее полураспада, то есть временем, необходимым для разрушения половины объема пены. Оба показателя зависят от рН среды, концентрации белка, солей, температуры, присутствия липидов, сахарозы, пищевых волокон, фракционного состава и строения белков. Для качества некоторых пищевых продуктов большое значение имеет размер пузырьков пен, который также зависит от технологических и других факторов. Глютенин пшеницы, например, образует пузырьки пены с большим размером, чем глиадин. После расщепления дисульфидных связей в глиадине и целой клейковине размер пузырьков не изменяется, в то время как у глютенина он уменьшается. На основе жироэмульгирующих свойств растительные и животные белки применяются в производстве хлебобулочных, мучных кондитерских изделий, низкокалорийных маргаринов, майонезов, паст, мясных продуктов, а пенообразующие свойства являются основой производства сбивных кондитерских изделий (бисквитов, десертов, кремов и т. д.). Способность белковых суспензий к сцеплению с поверхностями металла, пластмасс, картона, бумаги (адгезия) важна в процессах транспортировки, обработки, формования и упаковки тестовых, творожных, сырных, конфетных масс, мясных и рыбных фаршей, текстуратов белка и новых форм белковой пищи (аналогов). Гелеобразующие свойства белков характеризуются способностью их коллоидного раствора из свободно диспергированного состояния переходить в связнодисперсное (с образованием систем, обладающих свойствами твердых тел). Упругие свойства геля, обусловленные образованием пространственной сетки взаимодействующих молекул белка, зависят от минимальной его концентрации, при которой наступает гелеобразование (гель-точки), от рН, от присутствия других белков, солей, полисахаридов. Белок как гелеобразователь должен образовывать гели в широком диапазоне рН, ионной силы, при минимальной концентрации и с необходимыми физико-химическими свойствами. К последним относятся прочность, твердость, эластичность, тиксотропия (способность обратимо переходить в текучее состояние при механической обработке и вновь образовывать нетекучий гель после снятия нагрузки), температура размягчения и плавления, степень набухания, способность к синерезису (отделение дисперсионной среды с сокращением объема геля), сорбция красителей и ароматических веществ и т.д. К подобного рода "универсальным" гелеобразователям относится желатин, позволяющий в широких пределах обеспечить регулирование химического состава и биологическую ценность пищевых продуктов. Различают наполненные, смешанные, комплексные, анизотропные гели и ксерогели. Наполненные гели содержат другие белки в суспензированном или растворенном виде, смешанные состоят из пространственных сеток с разными видами белков, у комплексных гелей роль гелеобразователя выполняют комплексы белков с другими соединениями. Отличительной особенностью анизотропных гелей является наличие в их составе ориентированных молекул белка, а ксерогелей (сухих гелей) – возможность хранения их в течение длительного времени. Вязко-эластично-упругие свойства. Отличительным свойством некоторых пищевых белков является низкий уровень полярности функциональных групп. Молекулы воды, окружая частицы белков, отталкиваются, а молекулы белков, наоборот, агрегируются с образованием комплексов с присущими им реологическими свойствами (вязкость, эластичность, упругость). Наиболее выраженным комплексом таких свойств обладают белки пшеничной клейковины, обуславливающие текстуру хлеба и создающие непрерывную фазу в изделиях с наполнителями (зерно, отруби, изюм). За свойство упругости и эластичности белков ответственность несет глютениновая фракция белков. С целью обеспечения стабильности технологического процесса, улучшения качества и расширения ассортимента пищевых изделий осуществляют регулирование функциональных свойств. Функциональные свойства белков определяются их структурой. Например, вязкость и гелеобразующие свойства соотносятся с размером и формой молекул, а водосвязывающая способность, пенообразующие и эмульгирующие свойства коррелируют с соотношением на поверхности полярных и гидрофобных групп. Все факторы, которые изменяют структуру белков, вызывают и регулирование (модификацию) их свойств. Регулирование функциональных свойств белков достигается изменением условий их выделения, сушки, физическими, физико-химическими воздействиями, ферментативной и химической модификацией. Параметры обработки могут изменять аминокислотный и фракционный состав белков, вызывать денатурацию, агрегацию или взаимодействие с другими компонентами (липидами, углеводами). Наиболее широко используемыми методами регулирования функциональных свойств являются физико-химические и ферментативные. К физико-химическим методам относятся перевод белков перед сушкой в раствор кислот, щелочей, оснований – с целью изменения заряда или ионного состава, тепловая денатурация и т.д. При этом у белков улучшаются функциональные свойства: повышается растворимость, гелеобразующая, жироэмульгирующая способность, способность к текстурированию и прядению. Функциональные свойства белков улучшаются и за счет обработки их веществами липидной (лецитин, стеароил-2-лактилат натрия или кальция, моно- и диацилглицерины), углеводной (пектины, альгинаты, каррагинаны, камеди) или иной природы (поливалентные металлы). Реакционные группы белков взаимодействуют с различными типами соединений с образованием при этом композитных формул. Последние усиливают процессы водопоглощения, эмульгирования жира, гелеобразования, структурирования и тем самым улучшают качество готовых изделий. Хорошо изучено взаимодействие белков с заряженными полисахаридами, приводящее к получению нерастворимых электростатических комплексов. При этом наблюдается фазовое расслоение системы. Регулирование функциональных свойств белков в составе суспензии достигается добавлением в состав пищевых дисперсных систем, например мясных фаршей, анионных полисахаридов (пектаты, альгинаты, карбоксиметилцеллюлоза) или применением последних на стадии осаждения белка из растворов. Комплексообразование белков с полисахаридами эффективно и для избирательного фракционирования белковых компонентов за счет сорбции их как при одноименных, так и разноименных зарядах с молекулами анионов. Комплексы белок – анионный полисахарид имеют большую набухаемость, хорошую водоудерживающую способность и более высокие поверхностно-активные свойства. В присутствии, например, пектина и каррагинана эмульсионная емкость и стабильность жировой эмульсии с казеинатом натрия повышаются (Э. Токаев, И. Рогов и др., 1982). Образование комплекса белок – анионный полисахарид (КМЦ, гуммиарабик и др.) с казеином, глобулинами сои обеспечивает и получение более стабильных пен при более высокой массовой доле белка в них (В. Толстогузов, Е. Браудо и др., 1985). При образовании комплексов белка с анионными полисахаридами большая роль отводится взаимодействию положительно заряженных групп белка с отрицательно заряженными группами полисахаридов, а также образованию гидрофобных взаимодействий и водородных связей между комплексообразователями. При формировании таких комплексов иногда изменяется вторичная структура белка. Ферментативная модификация функциональных свойств белков осуществляется с использованием ферментов (табл. 2.14) растительного, микробного или животного происхождения. Преимуществом таких методов являются мягкие режимы выделения белков, сохранение биологической ценности и возможность регулирования глубины той или иной реакции. К недостаткам методов относится ограничение процессов модификации из-за высокой степени специфичности ферментов. Таблица 2.14.Методы ферментативной модификации белков [К. Швенке, 2000]
Из ферментативных методов модификации наибольшее распространение получил метод ограниченного ферментативного протеолиза. С его использованием для замены яичного альбумина выпускают пенообразующие модифицированные белки: гидролизаты изолята и гидролизаты муки. При ограниченном протеолизе, например, легумина кормовых бобов под влиянием трипсина – фермента пищеварительного тракта – расщепляются пептидные связи только α-цепей, тогда как (β-цепи остаются незатронутыми. И только в ходе дальнейшего глубокого протеолиза β-цепи 11S белка расщепляются на пептиды. Гидролитический распад гидрофильных а-цепей приводит к понижению молекулярной массы с 340 до 240 кДа и к возрастанию степени гидрофобности "Обрубка" – легумина Т. Молекулы белка становятся более компактными, сферическими и с высокой термодинамической стабильностью (Браудо Е. Е., 1997). При этом улучшаются эмульгирующие и пенообразующие свойства белков. Аналогичная взаимосвязь между особенностями структуры 11S глобулинов, подвергнувшихся ограниченному протеолизу, и функциональными свойствами существует и у гороха и у сои. Разница заключается в молекулярных массах легумина Т – 230–260 кДа. Определенный интерес представляют реакции ферментативного синтеза белков из пептидов (пластеиновый синтез), которые целесообразно применять для введения в состав белков незаменимых аминокислот или их производных (эфиров) с целью улучшения растворимости, поверхностно-активных свойств и биологической ценности:
Важнейшие химические методы модификации функциональных свойств приведены в табл. 2.15. Из этих методов широко известны приемы Таблица 2.15.Методы химической модификации белков [К. Д. Швенке, 2000]
* Продукты реакции нестабильны. дезамидирования (удаление амидных групп глютамина и аспарагина), ацилирования аминогрупп янтарным (сукцинилирование) или уксусным (ацетилирование) ангидридами и фосфорилирования. Сукцинилирование и ацетилирование протекают по следующим схемам:
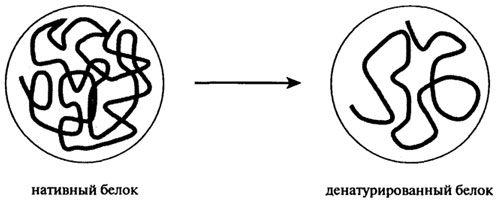
Данный вид химической модификации приводит к повышению суммарного отрицательного заряда молекулы из-за ковалентного присоединения остатков янтарной и уксусной кислот к ε-группам остатков лизина. Степень ацилирования возрастает за счет гидроксильных групп серина, треонина и тирозина. Электростатическое отталкивание одноименно заряженных групп приводит к структурным изменениям в белках и даже распаду 11S белков на субъединицы и развертыванию их глобулярной структуры. Подобные структурные изменения характерны для 11S глобулинов семян арахиса, рапса, подсолнечника, гороха и кормовых бобов. Благодаря изменению пространственной структуры и заряда молекул белков усиливаются гидрофобные свойства, следовательно, улучшаются эмульгирующие и пенообразующие свойства, изменяются растворимость и гелеобразующие свойства. Полипептидные цепи формируют гели при меньших значениях концентрации, рН и температуры, чем нативные белковые глобулы. С возрастанием степени модификации прочность гелей уменьшается, поэтому для ацетилированных белков целесообразна средняя степень модификации. Сукцинилированные или ацетилированные легумины при определенной степени модификации образуют эмульсии "масло в воде" высокой степени устойчивости. Фосфорилирование растительных белков с применением хлорокиси фосфора приводит к улучшению растворимости, эмульгирующих и пенообразующих свойств и способности к гелеобразованию. Стабилизации структуры геля способствуют ковалентные сшивки модифицированных белков: Белок-NНРОСl2 + НООС-Белок + 2Н2О → Бeлoк – NHCO – Бeлoк + 2НСl + Н3РО4 91 :: 92 :: 93 :: 94 :: 95 :: 96 :: 97 :: 98 :: 99 :: 100 :: 101 :: Содержание 101 :: 102 :: 103 :: 104 :: 105 :: 106 :: 107 :: 108 :: 109 :: 110 :: 111 :: 112 :: Содержание 2.9. ПРЕВРАЩЕНИЯ БЕЛКОВ Нативная трехмерная структура белков поддерживается разнообразием внутри- и межмолекулярных сил и поперечных связей. Любое изменение условий среды в технологических потоках производства пищевых продуктов оказывает влияние на нековалентные связи молекулярной структуры и приводит к разрушению четвертичной, вторичной и третичной структуры. Разрушение нативной структуры, сопровождающееся потерей биологической активности (ферментативной, гормональной), называют денатурацией. С физической точки зрения денатурацию рассматривают как разупорядочение конформации полипептидной цепи без изменения первичной структуры. Процесс денатурации протомера схематически представлен на рис. 2.17. Денатурация олигомерного белка заключается в диссоциации на протомеры, сопровождающейся или не сопровождающейся изменением их конформации.
Большинство белков денатурируются в присутствии сильных минеральных кислот или оснований, при нагревании, охлаждении, обработке поверхностно-активными веществами (додецилсульфатом), мочевиной, гуанидином, тяжелыми металлами (Ag, Pb, Hg) или органическими растворителями (этанолом, метанолом, ацетоном). Широкое применение кислот, оснований, солей, органических растворителей предусматривается в практике выделения белков из пищевого сырья и готовых продуктов при изучении их свойств и структурных особенностей, а также при экстракции и очистке в технологии выделения концентратов и изолятов. Денатурированные белки обычно менее растворимы в воде, так как их полипептидные цепи настолько сильно переплетены между собой, что затрудняется доступ молекул растворителя к радикалам остатков аминокислот. Большая часть белков денатурируется при 60–80°С, однако встречаются белки и термостабильные, например, α-лактоглобулин молока и α-амилазы некоторых бактерий. Повышенная устойчивость белков к нагреванию часто обуславливается наличием в их составе большого количества дисульфидных связей. Однако степень денатурирующего воздействия температуры на белки зависит и от их влажности, реакции и солевого состава среды и присутствия небелковых соединений. Например, температура денатурации белков сои и подсолнечника существенно понижается в присутствии кислот жирного ряда, в кислой и влажной среде, но повышается в присутствии сахарозы и крахмала. Факторы, вызывающие денатурацию белков, имеют особо важное значение для регулирования активности ферментов. Любые воздействия, направленные на стабилизацию вторичной и третичной структуры, приводят к повышению активности ферментов, а те, которые разрушают нативную структуру, – к их инактивации. При температуре от 40–60°С до 100°С со значительной скоростью протекает взаимодействие белков с восстанавливающими сахарами, сопровождающееся образованием карбонильных соединений и темноокрашенных продуктов – меланоидинов (реакция Майяра). Сущность реакций меланоидинообразования заключается во взаимодействии группы –NH2 аминокислот с гликозидными гидроксилами сахаров (см. гл. 3). Сахаро-аминные реакции являются причиной не только потемнения пищевых продуктов, но и уменьшения в них сухого вещества и потерь незаменимых аминокислот (лизина, треонина). Меланоидины понижают биологическую ценность изделий, так как снижается усвояемость аминокислот из-за того, что сахароаминные комплексы не подвергаются гидролизу ферментами пищеварительного тракта. К тому же количество незаменимых аминокислот уменьшается. Это уменьшение происходит не только за счет взаимодействия их с восстанавливающими сахарами, но и за счет взаимодействия между собой функциональных групп –NH2 и –СООН самого белка. Реакции протекают с образованием внутренних ангидридов, циклических амидов и ω–ε изопептидных связей. Механизм образования связей с участием глутаминовой кислоты и ее амида представлен на рис. 2.18. Изопептиды обнаружены в кератине, молочных белках и белках мяса.
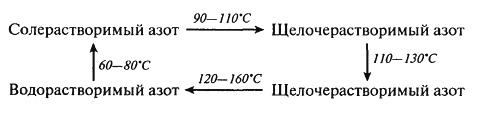
Тепловая денатурация белков является одним из основных физико-химических процессов, лежащих в основе выпечки хлеба, печенья, бисквитов, пирожных, сухарей, сушки макаронных изделий, получения экструдатов и сухих завтраков, варки, жарения овощей, рыбы, мяса, консервирования, пастеризации и стерилизации молока. Данный вид превращений относится к полезным, так как он ускоряет переваривание белков в желудочно-кишечном тракте человека (облегчая доступ к ним протеолитических ферментов) и обуславливает потребительские свойства пищевых продуктов (текстуру, внешний вид, органолептические свойства). В связи с тем, что степень денатурации белков может быть различной (от незначительной до полного изменения расположения пептидных цепей с образованием новых ковалентных –S–S-связей), то и усвояемость полимеров может не только улучшаться, но и ухудшаться. Параллельно могут изменяться физико-химические свойства белков. Для хлопковых семян, подвергнутых влаготепловой обработке, зафиксирован, например, переход растворимого азота из одной формы в другую:
Термическая обработка белоксодержащей пищи при 100– 120°С приводит не к денатурации, а к разрушению (деструкции) макромолекул белка с отщеплением функциональных групп, расщеплением пептидных связей и образованием сероводорода, аммиака, углекислого газа и ряда более сложных соединений небелковой природы. Так, стерилизация молочных, мясных и рыбных продуктов при температуре выше 115°С вызывает разрушение цистеиновых остатков с отщеплением сероводорода, диметилсульфида и цистеиновой кислоты:
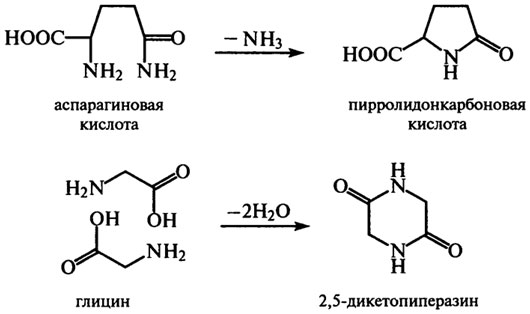
Реакции дезамидирования аспарагиновой и глутаминовой аминокислот и дегидратации глицина могут быть причиной образования новых ковалентных связей в белках, так как образуется пирролидонкарбоновая кислота и 2,5-дикетопиперазин (дикетопиперазина много в обжаренных бобах какао):
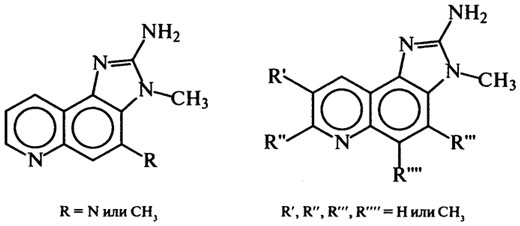
Среди продуктов термического распада белков встречаются соединения, придающие им мутагенные свойства. Термически индуцированные мутагены образуются в белоксодержащей пище в процессе ее обжаривания в масле, выпечки, копчения в дыму и сушки. Мутагены содержатся в бульонах, жареной говядине, свинине, домашней птице, жареных яйцах, копченой и вяленой рыбе. Некоторые из них вызывают наследственные изменения в ДНК, и их воздействие на здоровье человека может быть от незначительного до летального. В экстрактах, выделенных из жареной рыбы и мяса, еще в 70-х гг. найдены продукты пирролиза аминокислот, образующиеся обычно при 500–600°С. Продукты идентифицированы как Трп–П–1 и Трп–П–2 из триптофана, Фен–П–1 из фенилаланина, Глу–П–1 и Глу–П–2 из глутаминовой кислоты, Лиз–П–1 из лизина, Орн–П–1 из орнитина:
Другая группа мутагенных соединений в белковой пище открыта в 80-х гг. в умеренно нагретом мясе (ниже 200°С) и пищевых бульонах:
Токсические свойства белков при термической обработке выше 200°С или при более низких температурах, но в щелочной среде, могут обуславливаться не только процессами деструкции, но и реакциями изомеризации остатков аминокислот из L- в D-форму. Присутствие D-изомеров понижает усвояемость белков. Например, термообработка казеина молока при температуре около 200°С снижает биологическую ценность продукта на 50%. В сильнощелочных средах, особенно при высоких температурах, некоторые остатки аминокислот претерпевают ряд специфических превращений. Так, аргинин превращается в орнитин, цитруллин, мочевину и аммиак, а цистеин – в дегидроаланин с выделением сероводорода:
Реакционноспособный дегидроаланин конденсируется с остатками лизина, орнитина и цистеина боковых цепей и образует межмолекулярные поперечные связи в белках:
В реакцию конденсации могут вступать остатки аргинина, гистидина, треонина, серина, тирозина и триптофана. Питательная ценность белков с новыми поперечными связями ниже, чем у белков с нативной структурой, поэтому образование их в технологических процессах производства пищевых продуктов нежелательно. К тому же в опытах на крысах показано, что образование, например, лизиноаланина стимулирует нефрокальциноз, диарею и облысение. Обработка сырья растворами щелочей широко используется при получении изолятов и концентратов белков. Чем ниже значение рН, температура и время обработки, тем выше содержание незаменимых аминокислот в белке. Например, при повышении рН раствора с 8,5 до 12,5 при экстракции белка из пшеничных отрубей количество лизина в нем уменьшается на 40%, треонина – на 26%, а валина – на 24%. Мягкие температурные режимы предохраняют от образования в больших количествах нежелательных аминокислотных фрагментов. В то же время среди специалистов обсуждается вопрос о введении предельно допустимых концентраций лизиноаланина (например, 300 мг на 1 кг) в целях обеспечения безопасности белоксодержащей пищи. Неблагоприятные погодные условия при созревании зерна, поражение его вредителями и микрофлорой, пониженная или повышенная температура при хранении и переработке сырья и полуфабрикатов, механические, физические и химические факторы (перемешивание, гомогенизация, замес, инфракрасное облучение, ультразвук, действие солей, диоксида углерода, газообразных азота и этилена и т. д.) усиливают структурные перестройки белков, которые могут разрушать природные белково-липидные взаимодействия. Высвобождающиеся липиды, подвергаясь окислительной порче, способны инициировать образование ковалентных меж- и внутримолекулярных связей в белках и новых полимерах:
Остатки тирозина в присутствии гидропероксидов (LO2H) могут превращаться в сульфоксиды и сульфоны, остатки цистеина – в сульфиновые, сульфоновые кислоты, а остатки триптофана – в гидрокси-β-индолилаланин и N-формилкинуренин. Все реакции окисления связаны с потерей незаменимых аминокислот. Окислительная порча белков особенно опасна при переработке масличного и жирового сырья. Торможение реакций можно достигать добавлением оксидантов, ферментных препаратов или повышением активности собственных ферментов сырья в целях вывода липидов из взаимодействия с белками. Использование новых и традиционных технологических процессов без глубокого изучения влияния их на молекулярные основы структуры белков, с одной стороны, опасно для здоровья людей, а с другой – неэффективно с точки зрения обеспечения качества пищевых продуктов. Примером может служить научно обоснованное применение аскорбиновой кислоты для улучшения качества хлеба. По современным представлениям, аскорбиновая кислота, окисляясь кислородом воздуха в дегидроаскорбиновую кислоту, окисляет глютатион (Г-SH), превращая его в окисленную форму (Г-S-S-Г). Это предохраняет глютатион от участия в сульфгидрильно-дисульфидном взаимодействии с белками клейковины во время образования теста. Сульфгидрильные группы пшеничного белка взаимодействуют друг с другом с образованием дисульфидносвязанных белковых цепей и качество изделий улучшается. Наряду с окислительными процессами в технологических процессах, предусматривающих механические или физические воздействия на белковые вещества сырья (замес, гомогенизация, ультразвук и т.д.), протекают и другие превращения, характер которых зависит от природы, степени и способа этих воздействий. На начальных стадиях замеса хлебного теста и при измельчении семян зерна наблюдается тепловая агрегация белков, при усиленной механической обработке теста возможна деструкция последних с разрывом дисульфидных и даже пептидных связей. Агрегирующая и комплексообразующая способность белков пшеницы является одним из важных показателей, обеспечивающих им ведущую роль в формировании клейковины в процессе ее отмывания из муки и тестоведения. Параметры агрегации определяют по методу японских исследователей (Т. Arakawa, D. Yonezawa, 1975). Для этого смешивают растворы клейковинных белков в 0,01 н уксусной кислоте и 0,2 М натрий-фосфатного буферного раствора (рН 5,6), содержащего 2 М NaCl. Измеряют оптическую плотность раствора (λ = 350 нм) во времени агрегации процесса и рассчитывают коэффициент начального этапа агрегации (К) и показатель агрегации (t10/c), характеризующий степень помутнения раствора в течение 10 мин. Константу начальной скорости агрегации рассчитывают по уравнениям: К =
и τ3 = 3rt + τ03, где τ – оптическая плотность раствора при λ = 350 нм; r – постоянная, отражающая изменение τ3 в первые 1–1,5 мин агрегации; t – время агрегации; τ0 – оптическая плотность при t = 0; с – концентрация белка, %. Параметры агрегации белков сильных пшениц, характеризующиеся более "плотной" пространственной упаковкой структуры, выше по сравнению со слабыми, имеющими более "рыхлую" организацию молекул (рис. 2. 19). Процесс образования белковых агрегатов по ходу технологического процесса приготовления изделий из муки интенсивнее у крепкой клейковины, чем у слабой.
Константы агрегации уксуснорастворимых белков, экстрагированных из предварительно отмытой клейковины и теста, выше, чем белков, выделенных из муки [В. Колпакова, Е. Молчанова, 1991]. Следовательно, при гидратации белков в ходе технологического процесса и, в частности, при замесе теста или отмывании клейковины водой происходят внутри- и(или) межмолекулярные превращения, связанные с изменением структуры молекул, приводящим к снижению агрегирующих свойств последних. Наиболее высокие показатели агрегации наблюдаются у α- и γ-компонентов глиадина, наименее – у ω-компонентов. Способность к агрегации возрастает по мере увеличения подвижности полипептидов и уменьшения их молекулярной массы. Очень высокой способностью образовывать надмолекулярные ассоциаты обладают быстроподвижные α-глиадины с молекулярной массой 31 000. Агрегирующая способность белков взаимосвязана с особенностями аминокислотного состава. Так, суммарный глиадин с более повышенной агрегирующей способностью содержит меньше заряженных групп, а ω-глиадины, обладающие низкой степенью агрегации, богаты пролином, фенилаланином, амидогруппами и полярными аминокислотными остатками. Это определяет высокую способность их к взаимодействию с другими компонентами муки и, прежде всего с липидами и липидоподобными соединениями (дигалактозилдиглицерид). Комплексообразование последнего с ω-глиадинами за счет водородных связей обуславливает газоудерживающую способность теста и объем хлеба. Имеются данные, что структура комплексов белки–липиды имеет сходство со структурой белок-липидных мембран. В агрегации и взаимодействии молекул клейковинных частиц с другими компонентами муки в технологических процессах производства хлеба, макаронных изделий, печенья и т.д., наряду с водородными связями, принимают участие гидрофобные, ионно-электростатические и дисульфидные связи. Способность белков к формированию высоко агрегированных и надмолекулярных образований зависит от рН, ионной силы и состава среды (присутствие денатурирующих, солюбилизирующих, восстанавливающих и других агентов). С повышением рН от 4,0 до 9,1 агрегация белков злаковых культур (пшеницы, ржи, ячменя) повышается. Чем больше концентрация нейтральных солей, тем агрегирующая способность белков выше. Отсюда, определенные дозировки поваренной соли способны регулировать образование или разрушение макромолекул клейковинного белка в состоянии пространственно организованной структуры пшеничного теста при производстве, например, хлеба. Определенное влияние на агрегативное состояние запасных белков оказывают детергенты (ПАВ) различной природы: анионные, катионные, нейтральные. В присутствии анионного детергента в кислой среде, когда молекулы имеют положительный заряд, агрегация белка максимальна. Действие же катионных и нейтральных детергентов в направлении усиления или ослабления агрегации неоднозначно и зависит от рН среды и их концентрации. В целом же, агрегирующая способность запасных белков различных зерновых культур в ходе технологического процесса может быть представлена в виде интегральной суммы множества взаимодействий, способствующих или препятствующих образованию надмолекулярных структур: ∑A = ∆Аи +∆Аг+∆Ав +∆Ан +∆Аα +∆Ад + ..., где ∆Аи – агрегация белка за счет ионно-электростатических сил; ∆Аг – гидрофобных взаимодействий; ∆Ав – водородных связей; ∆Ан – набухания (гидратации); ∆Аα – изменения регулярных структур; ∆Ад – содержания дисульфидных связей. Степень участия того или иного фактора в изменении агрегативного состояния белков, а с ним и реологических свойств теста и качества хлеба, зависит от исходных физико-химических свойств и структурных особенностей белков муки, технологических факторов процесса (температура, степень механического воздействия, рН среды и т. д.), химической природы и количества дополнительного сырья, улучшителеи и пищевых добавок. Огромное значение для изменения свойств и агрегативного состояния белков в ходе технологического процесса приготовления теста имеет дисульфидно-сульфгидрильный обмен: P1S-SP2 + P3SH → Р1S – SР3 + P2SH Считают, что лабильность дисульфидных связей обеспечивает образование новых прочных межмолекулярных связей и релаксацию (ослабление) напряжения структур клейковины при замесе теста. Сильная мука, содержащая крепкую клейковину, требует больше времени замеса, так как она содержит больше дисульфидных связей и меньше SH-групп, чем слабая. В процессе созревания муки при ее отлежке, а также под влиянием окислителей типа броматов, улучшение реологических свойств клейковины объясняется увеличением содержания S-S-связей и уменьшением, соответственно, SH-групп. В присутствии восстанавливающих агентов (сульфит натрия) реологические свойства клейковины ухудшаются, как это имеет место в прорастающем зерне или в процессе приготовления теста из зерна, пораженного клопом-черепашкой. Во всех случаях показатели качества и реологические характеристики теста взаимосвязаны с изменением в соотношении S–S-связей и SH-групп в белках. Процессы образования или распада S– S-связей в созревающем и прорастающем зерне пшеницы протекают при участии специфических ферментов тиол–дисульфидного обмена: тиол– кислородоксидоредуктаза (КФ 1.8.3.2) и тиол–протеиндисульфидредуктаза (КФ 1.8.4.2). Предполагают, что направленное регулирование активности указанных выше ферментов в процессе приготовления теста является действенным приемом улучшения технологических свойств муки в целях обеспечения надлежащего качества хлеба [Т. Горпинченко, 1995]. В результате действия протеолитических ферментов в технологическом потоке производства пищевых продуктов белковые вещества так же претерпевают ряд существенных изменений. Так, на стадии солодоращения при производстве пива в эндосперме ячменя наблюдается гидролиз глобулина (эдестин), альбумина (лейкозин), проламина (гордеин) и глютелина с накоплением азотистых соединений с более низкой молекулярной массой (пептиды, аминокислоты). В результате в зерне накапливается растворимая, коагулируемая и аминная формы азота, тогда как в зародышевом листке и ростках зерна, наоборот, увеличивается количество белкового азота за счет процессов синтеза. Высокомолекулярные фракции белков при прорастании зерна расщепляются под действием ферментов эндопептидаз по схеме:
и экзопептидаз (дипептидаз и полипептидаз). Полипептидазы включают аминопептидазы (а-аминоацилпептидгидролазы, 3.4. 1 ), для действия которых необходимы свободные аминогруппы (– NH2), и карбоксипептидазы (пептидиламинокислотные гидролазы, 3.4.2), требующие для гидролиза свободные карбоксильные группы (– СООН). Технологические режимы проращивания зерна предусматривают оптимальные условия (рН, температура) действия гидролитических ферментов для обеспечения накопления низкомолекулярных азотистых соединений для питания дрожжей сусла. В традиционной технологии производства хлеба и мучных кондитерских изделий из муки нормального качества при тестоведении не протекают глубокие реакции гидролиза белков, свидетельствующие об изменении их первичной структуры, тогда как слабые процессы пептизации низкомолекулярных азотистых соединений, являющихся продуктами незавершенного синтеза в зерне, могут иметь место. Изменения структуры белков в процессе тестоприготовления ограничиваются, как правило, дезагрегацией, агрегацией молекул и изменением высших уровней ее организаций (вторичная, третичная, четвертичная структуры). 101 :: 102 :: 103 :: 104 :: 105 :: 106 :: 107 :: 108 :: 109 :: 110 :: 111 :: 112 :: Содержание 112 :: 113 :: 114 :: 115 :: 116 :: 117 :: 118 :: 119 :: 120 :: 121 :: Содержание 2.10. КАЧЕСТВЕННОЕ И КОЛИЧЕСТВЕННОЕ Присутствие белков в пищевых объектах устанавливается с помощью качественных реакций, которые условно разделяют на две группы: а) цветные реакции; б) реакции осаждения. Среди первой группы различают универсальные реакции (биуретовая на пептидные связи и нингидриновая на α-аминокислоты) и специфические, обусловленные присутствием в белках остатков определенных аминокислот. Так, ксантопротеиновая реакция свидетельствует о наличии в белках остатков ароматических аминокислот, реакция Паули – гистидина и тирозина, Адамкевича и Вуазене – триптофана, нитропруссидная – цистеина, а реакция Сакагучи – аргинина. По результатам специфических реакций ориентировочно можно судить о пищевой ценности белков. Во второй группе реакций белки осаждают действием солей, органических растворителей, концентрированных кислот, щелочей, ионов тяжелых металлов, температуры и в изоэлектрической точке. Белки в растворенном состоянии крайне неустойчивы, поэтому при добавлении органических растворителей (спирт, ацетон), концентрированных растворов нейтральных солей щелочных металлов и воздействии физических факторов (нагревание, облучение, ультразвук) гидратная оболочка разрушается и они выпадают в осадок. Так как белковые вещества сырья (муки, крупы, молока, мяса), включая ферменты, часто являются определяющими в обеспечении качества пищевых изделий, то для изучения физико-химических, биохимических и физиологических свойств этих соединений обязательным условием является получение белков в индивидуальном и, по возможности, неденатурированном состоянии. Белки обычно теряют природные (нативные) свойства (растворимость, гидратацию, ферментативную активность и т.д.), подвергаясь денатурации под влиянием различных факторов. Типичным примером необратимой денатурации белков является выпадение их в осадок под действием ТХУ. Длительный контакт со спиртом также приводит к необратимой денатурации белка. Денатурирующее действие различных факторов на белки можно смягчить, если проводить операции выделения их при температуре не выше +4°С. Методы выделения и очистки белков. Общая схема операций по выделению белков сводится к измельчению биологического материала (гомогенизации), экстрагированию и собственно выделению, то есть очистке и получению белка в индивидуальном состоянии. При изучении метаболических процессов в живых организмах (в созревшем зерне, плодах, овощах) морфологическая и биохимическая целостность клеток и тканей сохраняется в максимальной степени, тогда как при исследовании состава сырья и готовых пищевых продуктов потеря целостности структуры несущественна. Гомогенизацию объектов следует рассматривать как начальную стадию выделения белков, но способ ее определяется постановкой задачи. Например, анализ ферментов из растительных материалов часто затруднен тем, что при гомогенизации экстрагируется большое количество фенолов, которые взаимодействуют с карбонильными группами пептидных групп при помощи водородных связей и вызывают денатурацию белка и потерю ферментами своей активности. Добавление в экстракт поливинилпирролидона, образующего с фенолами нерастворимые комплексы, предотвращает инактивацию ферментов. Разрушение клеточной структуры осуществляется тщательным измельчением материала в гомогенизаторах, мельницах, попеременным замораживанием и оттаиванием, применением ультразвуковых высокочастотных колебаний, пресс-методов с использованием высоких давлений и метода "азотной бомбы". В последнем случае клетки насыщаются азотом под давлением, которое затем сбрасывается и клетки разрушаются. Эффективность гомогенизации зависит не только от способа разрушения клеточных структур, но и от вида анализируемого материала. Животные клетки разрушаются относительно легко, особенно в отсутствие сосудистой и соединительной ткани, тогда как растительные и микробные – из-за присутствия клеточных стенок – трудно. В таком случае применяют методы растирания материала с твердыми веществами (песок, абразивный порошок) или обработку клеточных стенок лизоцимом или ферментными препаратами, содержащими целлюлазу, хитиназу и липазу. Гомогенизацию рекомендуется проводить в холодных комнатах или с использованием льда. Экстракция белков может быть совмещена с гомогенизацией клеток и тканей либо проведена отдельно, если продукт заранее измельчен. Для определения ферментативной активности белка достаточно одноразовой экстракции, тогда как для количественного определения белковых фракций зерна – трех- или пятикратной. Условия экстрагирования белков (время, гидромодуль, температура и т.д.) подбираются эмпирически, основываясь на методиках ведущих научных школ. Большинство белков животных тканей хорошо растворимы в 5–10% растворах солей, тогда как для перевода в раствор белков зерновых культур применяют более широкий набор растворителей. Для этого используются буферные системы со значениями рН от кислых до слабощелочных (фосфатные, боратные, цитратные, трис-HCl), органические растворители и неионные детергенты, разрывающие белок-липидные или белок-белковые связи:
Растворители подбираются с учетом разрыва в белках определенных типов связей. Так, уксусная кислота ослабляет ионные связи, сообщая молекулам одноименные положительные заряды, мочевина – водородные и гидрофобные, салицилат натрия и ДДС-Na – гидрофобные и ионные, а водные растворы спиртов – водородные и гидрофобные взаимодействия. Органические растворители разрывают белок-липидные связи. При изучении физико-химических свойств белков и их превращений в пищевых системах широко используют методы фракционирования и очистки от небелковых соединений. Они основаны на различиях таких свойств белков, как размер молекул, растворимость, заряд и сродство к специфическим химическим группам. Осаждение белков из раствора под действием солей щелочных и щелочноземельных металлов называют высаливанием. Для высаливания чаще применяется сульфат аммония, под влиянием которого белки, как правило, сохраняют растворимость и ферментативную активность. Главную роль при высаливании играет не природа солей, а валентность ионов, действие которых оценивается по ионной силе (μ): μ = ½ ∑CV2, где С – концентрация каждого вида иона; V – валентность ионов. Глобулины выпадают в осадок при 50% насыщении, альбумины – при 100% насыщении растворов солей, а при ступенчатом фракционировании (20–100% насыщения) выпадают белки и других классов (проламины, глютелины). В практике выделения и очистки белков используются различные типы хроматографии: адсорбционная, распределительная, ионообменная и хроматография по сродству. Адсорбционная хроматография основана на различиях в полярности белков. В колонке вместе с буферным раствором упаковывают адсорбент, на который в небольшом объеме растворителя наносят исследуемый образец. Компоненты разделяемой смеси адсорбируются, затем элюируются с помощью буферного раствора с увеличивающейся концентрацией или полярностью. Фракции белка собирают с помощью автоматического коллектора фракций. В распределительной хроматографии, в отличие от адсорбционной, в качестве неподвижной фазы выступает водный слой, удерживаемый твердой фазой (силикагель, бумага). Разделяемые вещества многократно распределяются между водным слоем и движущейся фазой растворителя и с разной скоростью перемещаются подлине колонки или бумаге. Распределительную хроматографию на бумаге часто используют для анализа пептидов и аминокислот. Адсорбентом служат нити целлюлозы, а растворителем – смесь органических растворителей, например: бутиловый спирт–уксусная кислота–вода. Хроматограмму проявляют, высушивают и анализируют местонахождение разделяемых компонентов тем или иным способом. Методом ионообменной хроматографии белки или аминокислоты разделяют на основе различий в общем заряде молекул. Если белок в нейтральной среде (рН 7) имеет положительный заряд, то он связывается на колонке с ионообменником, содержащим фенольные, сульфо- и карбоксильные группы (катионообменник), если отрицательный, то – на колонке с ионообменником, представленным аминами или органическими основаниями (анионообменник). Чаще всего для фракционирования белков используют производные полистирола и целлюлозы:
Положительно заряженный белок снимается с колонки с помощью раствора хлористого натрия или изменением рН элюирующего буфера. При этом ионы натрия конкурируют с положительно заряженными группами белков. Белки с меньшим положительным зарядом вымываются с колонки первыми, с большим зарядом – последними. Хроматография по сродству (аффинная хроматография,) основана на принципе избирательного связывания белков со специфическими веществами (лигандами), прикрепленными к носителю. Лиганд (глюкозу) ковалентно присоединяют к носителю (проводят иммобилизацию) и наносят на колонку исследуемую белковую смесь. Несвязавшиеся белки удаляют соответствующим буфером, а нужный белок элюируют раствором, содержащим лиганд в очень высокой концентрации. При этом присоединенные к колонке остатки глюкозы в молекуле белка замещаются на глюкозу, находящуюся в растворе (рис. 2.20).
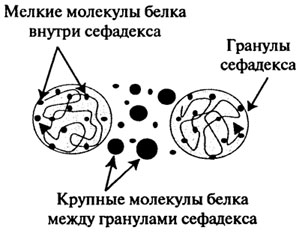
Гель-фильтрация, или метод молекулярных сит заключается в пропускании белков через колонку с гелем сефадекса или других типов (агарозных, полистирольных). Применяются также пористые стеклянные шарики и пористый кварц (порасил). Наибольшее распространение получили декстрановые гели (сефадекс), являющиеся продуктом поперечного сшивания полисахаридных цепочек декстрана. Зерна сефадексов разных номеров содержат поры разных размеров, в которые могут проникать белки с определенной молекулярной массой. Низкомолекулярные белки распределяются в растворенном виде как внутри частиц полимера, так и между ними, а высокомолекулярные – только между частицами, поэтому вторые быстрее проходят через колонку и первыми вытекают из нее (рис. 2.21). В итоге белки распределяются по молекулярной массе и могут быть собраны в виде отдельных хроматографических фракций (рис. 2.22).
Принцип методов электрофоретического разделения заключается в способности молекул пептидов и аминокислот, находясь в заряженной форме в виде катионов (+) или анионов (–), передвигаться в электрическом поле с определенной скоростью. Кроме того, молекулы с близкими зарядами, но разными размерами, отличаются отношением заряда к массе. Все эти различия и обуславливают высокую разрешающую способность электрофоретических методов. Скорость миграции белков в электрическом поле (V) зависит от напряжения электрического поля(ε), заряда белков (z) и сопротивления трения (О-Сопротивление трения определяется размерами, формой белка, значениями рН и концентрацией буфера. Указанные величины связаны между собой соотношением: V = ε · z / f. Впервые метод электрофореза был разработан Тизелиусом с применением бумаги в качестве носителя и специальных оптических устройств, регистрирующих передвижение границы раздела раствора белка и растворителя по показателям преломления (фронтальный электрофорез). В настоящее время распространены методы зонального электрофореза, предусматривающие использование крахмальных и полиакриламидных гелей (ПААГ). Наиболее распространенным методом фракционирования белков является диск-электрофорез (от англ, discontinuous – прерывистый) в ПААГ, при котором используется пара буферных растворов с различными значениями рН в присутствии ДДС-Na и гели различной пористости (концентрирующие и разделяющие) (Laemmli, 1970). Для обнаружения белков гели обрабатывают красителями: амидовым черным 10В, кумасси синим R-250. Интенсивность окраски, а по ней количественное содержание белковых фракций, определяют сканированием на денситометре. Для электрофоретического разделения белков и пептидов успешно применяется двумерный электрофорез в ПААГ. В соответствии с этим методом смесь компонентов разделяют сначала в столбиках геля электрофорезом в горизонтальном направлении, затем в гелевых пластинах – в вертикальном (рис. 2.23). При разделении белков, например гороха, этим методом удалось получить более 150 различных компонентов.
Очень высокую разрешающую способность имеет метод изоэлектрического фокусирования белков, в основе которого лежит фронтальный электрофорез, проводимый на колонке одновременно в градиенте рН и напряжения. Колонку предварительно заполняют носителями с синтетическими смесями полиаминополикарбоновых кислот (амфолитами), затем сверху в нее подают раствор сильной кислоты, снизу – сильнощелочной раствор для того, чтобы установить градиент рН с крайними значениями, соответствующими рН кислого и щелочного растворов. Амфолиты прекращают движение по колонке, когда их суммарный заряд становится равным нулю, и тем самым стабилизируют исходный градиент рН. В подготовленную колонку вносят образец исследуемой смеси, компоненты которой распределяются по зонам со значениями рН, характерными их изоэлектрическим точкам. В химии пищевого белка применяют и другие разновидности электрофоретическогоразделения(иммуноэлектрофорез, изотахофорез), а также метод пептидных карт и ультрацентрифугирование. Метод пептидных карт (отпечатков пальцев) относится к методам двумерного разделения и наиболее часто используется для анализа пептидов. Пептиды получают избирательным гидролизом белков, затем на бумаге их разделяют в горизонтальном направлении электрофорезом, в вертикальном – распределительной хроматографией. Пептиды окрашивают нингидрином, элюируют и определяют аминокислотный состав. В методе ультрафильтрации белки в градиенте плотности распределяются на разных уровнях центрифужной пробирки в процессе седиментации (осаждения) в виде отдельных зон. Для создания градиента используют соли тяжелых металлов и растворы сахарозы. Метод широко применяется для определения молекулярных масс белков по константе седиментации (S), которая зависит от массы и формы белковых частиц: S = v/(w2 · r), где v – скорость перемещения границы растворитель–белок, см/с; w– угловая скорость ротора, рад/с; г – расстояние от центра ротора до середины ячейки с раствором белка. Величина S, равная 1 · 10-13 с, принята за единицу и названа сведбергом (S) в честь Т. Сведберга, впервые сконструировавшего ультрацентрифугу. Очистка белков от низкомолекулярных соединений (солей, сахаров, аминокислот) осуществляется методами диализа, гель-фильтрации на сефадексе G-25, кристаллизации, ультрафильтрации и с помощью полых волокон. При диализе используют полупроницаемые мембраны (целлофан, коллодийная пленка), через которые белки не диффундируют и остаются внутри диализного мешочка. Более мелкие молекулы проходят через поры диализной мембраны и выходят в диализат. В методе ультрафильтрации, который применяется, например, в производстве сывороточных белков молока, соевых белковых изолятов, по обе стороны мембраны создается разность давлений за счет продавливания фильтруемого белкового раствора. В качестве мембран используются целлюлозные пленки и нецеллюлозные полиэлектролитные комплексы. Аналогично мембранам по принципу молекулярного сита действуют и полые волокна. Белковый раствор помещается с внешней стороны волокон, и создается разность давления за счет повышения его в растворе или понижения внутри их. Гомогенность белка определяется на последнем этапе выделения и очистки с применением по меньшей мере двух методов, оценивающих то или иное физико-химическое свойство. Наиболее достоверными являются ультрацентрифугирование в градиенте плотности, диск-электрофорез в ПААГ, иммунохимические методы и растворимость. Если белок при электрофорезе представлен только одной полосой и обладает при этом максимальной биологической активностью, то он считается гомогенным. Для гомогенного белка на кривой растворимости (зависимости растворенного белка от общего его количества в постоянном объеме растворителя) имеется только один перегиб, тогда как для гетерогенного – столько, сколько в нем индивидуальных компонентов. Содержание белка в пищевых объектах обычно определяют по количеству азота с использованием метода Кьельдаля. С целью упрощения и сокращения длительности анализа этот метод с момента его разработки (1983) неоднократно модифицировался с применением различных катализаторов и условий минерализации. На основе модифицированных методов созданы высокопроизводительные автоматические анализаторы типа "Кьельфос", стоимость определения содержания белка на которых и сегодня остается высокой. Существует и некоторая ус
Поиск по сайту: |