|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Тема 8. Генные мутацииСтр 1 из 2Следующая ⇒
Генные мутации – это наследственно закрепленные изменения генетического материала на уровне отдельных генов. Генные мутации используют для идентификации генов, картирования хромосом и генов, в биохимической генетике для установления функций гена, последовательности этапов биосинтетического пути, а также в селекционной практике. Кроме того, с помощью генных мутаций изучают механизмы спонтанного и индуцированного мутагенеза, оценивают специфичность действия мутагенов, исследуют роль репликации, репарации и рекомбинации в мутационном процессе. Генные мутации бывают прямые (приводящие к мутантному фенотипу) и реверсии (приводящие к дикому типу). Если после мутагенеза восстанавливается исходная пара нуклеотидов, мутация называется обратной или истинной реверсией, если же восстановление дикого типа обеспечивается мутацией в другом участке гена или за его пределами, то мутация носит название супрессорной (внутригенная или внегенная супрессия, соответственно). Возможность получения ревертантов служит важной характеристикой, позволяющей отличить замены пар оснований в гене от вставок или делеций. Изучение генных мутаций может осуществляться на трех уровнях: 1 – на молекулярном уровне (на уровне ДНК), 2 – на уровне белка, 3 – на уровне признака. Генные мутации могут быть спонтанные и индуцированные. Молекулярные механизмы возникновения мутаций. Спонтанные мутации возникают в процессе репликации ошибочных оснований, которые появляются в ДНК в результате повреждения оснований – апуринизации, дезаминирования или их окисления. Возникающие при этом мутации могут быть транзициями (замена пурина на пурин) или трансверсиями (замена пурина на пиримидин, или наоборот). В случае апуринизации нуклеотид теряет пуриновое основание (например, гуанин) вследствие разрыва фосфодиэфирной связи между основанием и остатком дезоксирибозы, что приводит к образованию «апуринового сайта», напротив которого при следующем акте репликации в синтезирующейся нити ДНК встраивается любой нуклеотид (чаще всего аденин), что в конечном итоге приводит к возникновению мутации. Например, пара ГЦ меняется на АТ (транзиция). Cпонтанному дезаминированию может подвергаться цитозин или 5-метилцитозин. В случае отщепления NН2-группы от цитозина, он превращается в урацил, и в ходе следующих актов репликации вместо пары ЦГ в ДНК появляется пара УА, а затем ТА. Если дезаминированию подвергается 5-метилцитозин, образуется тимин, который затем спаривается с аденином, в результате чего пара ЦГ заменяется на ТА (транзиция). Причиной возникновения спонтанных мутаций может быть также таутомеризация оснований. Известно, что азотистые основания могут находиться в четырех формах: кето-, и енольной (характерно для тимина и гуанина), амино- и иминоформе (характерно для цитозина и аденина). Кето- и аминоформа являются нормой, а переход оснований в енольную или иминоформу приводит к образованию неправильных пар и вызывать мутации типа замены пар оснований. Тимин в енольной форме в ходе репликации ошибочно спаривается с гуанином, гуанин в енольной форме – с тимином, цитозин в иминоформе – с аденином, аденин в иминоформе – с цитозином. Спонтанные мутации могут возникнуть также в результате случайной потери или вставки дополнительного нуклеотида в ходе репликации ДНК. Индуцируют появление мутации физические факторы: УФ-свет, g-лучи, СВЧ и химические агенты: нитрозогуанидин (НГ), нитрозометилмочевина, этилметансульфонат, HNO2, 5-бромурацил, 2-аминопурин, акридиновые красители и т.д.), а также транспозоны и некоторые вирусы. Радиация чаще всего вызывает разрывы молекулы ДНК, образующиеся либо в результате прямого действия лучей, либо опосредованно под действием возникающих в клетке радикалов. УФ-свет, в основном, индуцирует образование пиримидиновых димеров. Химические агенты действуют на ДНК различными способами: они могут вызывать дезаминирование, алкилирование оснований, что приводит к замене пар оснований, а также индуцировать выпадение и вставки отдельных нуклеотидов. В соответствии с этим, индуцированные мутации могут быть транзициями, трансверсиями, вставками и выпадениями нуклеотидов (табл. 11). При анализе генных мутаций на уровне ДНК необходимо помнить, что повреждение нуклеотидов может быть как в кодирующей, так и не кодирующей нити ДНК. В молекуле ДНК транскрибируется 3`®5` нить (матричная нить), в результате образуется 5`®3` нить мРНК, трансляция которой начинается на 5`-конце с АУГ кодона. В соответствии с этим матричная нить ДНК (3`®5`) называется смысловой, а комплементарная нить ДНК (5`®3`) – антисмысловой (рис. 12). Таблица 11 Типы мутаций, вызываемые мутагенами
Поскольку антисмысловая нить ДНК идентична синтезированной мРНК (за исключением того, что в ДНК – тимины, а в РНК – урацилы), она называется кодирующей, а смысловая, соответственно, некодирующей.
Рис. 12. Последовательность мРНК комплементарна матричной нити ДНК, с которой она транскрибируется Для того чтобы определить аминокислотную последовательность полипептида по нуклеотидной последовательности мРНК используют таблицу генетического кода (табл. 12). По характеру проявления на уровне белка генные мутации классифицируются как миссенс и нонсенс-типа. В случае возникновения миссенс мутации замена пары нуклеотидов приводит к замене аминокислоты в белке и снижению или утрате его функции. При возникновении мутации нонсенс типа замена нуклеотида в ДНК приводит к образованию бессмысленного кодона в мРНК (стоп-кодона) - УАГ, УАА или УГА, что вызывает преждевременную остановку трансляции и белок образуется абортивный. Добавление или выпадение одного или двух нуклеотидов вызывает мутации со сдвигом рамки считывания (frame-shift), в результате чего изменяется последовательность нуклеотидов в ДНК, а также смысл кодонов в мРНК, и, следовательно, правильное считывание полипептида в процессе трансляции. Таблица 12 Генетический код
На молекулярном уровне генные мутации могут проявляется в изменении последовательности нуклеотидов в ДНК, смысла считываемой мРНК, рРНК или тРНК, и, как результат этого, изменению аминокислотной последовательности синтезируемых полипептидов, либо полной остановке их синтеза. Анализ генных мутаций на уровне признака производится путем исследования их фенотипов. Генные мутации могут быть доминантными и рецессивными. Для некоторых генов характерно явление множественного аллелизма. В гетерозиготном состоянии генные мутации могут находиться в аллельных взаимодействиях – доминантно-рецессивном, неполном доминировании и кодоминировании. Иногда генные мутации в гомозиготном состоянии могут вызывать летальный эффект. Генные мутации наследуются согласно законам Г. Менделя (при отсутствии сцепления), сцеплено с полом (при локализации в Х- или У-хромосоме), или сцеплено (в зависимости от их расположения на хромосоме относительно друг друга). При экстрахромосомальной локализации генные мутации наследуются согласно принципам внехромосомного наследования. При проведении количественного учета генных мутаций пользуются двумя основными методами – определяют частоту появления мутаций, либо скорость мутирования. Частота мутаций определяется величиной, с которой мутация определенного типа обнаруживается в популяции клеток или целых организмов. Клеточной популяцией при этом могут быть гаметы, споры или любые другие типы клеток. Частота мутаций выражается формулой:
n – число мутантных клеток (организмов), а N – общее число анализируемых клеток (организмов).
Скорость мутирования характеризует частоту появления мутаций в гене или геноме за определенный промежуток времени. Эта величина определяется как число мутаций на ген (геном) на генерацию (клеточное деление) или поколение и равна: n – число мутантных клеток, N – общее число анализируемых клеток, а g – число генераций, которое прошло от первого деления клетки до момента проведения анализа. Число N может быть также выражено формулой: N = 2g. РЕШЕНИЕ ТИПОВЫХ ЗАДАЧ В этом разделе будет представлено решение 2-х типов задач: 1. Молекулярные механизмы возникновения мутаций. 2. Определение частоты мутаций. Задача 1.Вы изолировали белок из организма дикого типа и соответственно мутантный белок из мутантного организма и определили их аминокислотный состав. Вы обнаружили, что они отличаются только одной аминокислотой. Дикий тип: Arg Met Ser Мутант: Arg Ile Ser. Замена аминокислоты Met на Ile может произойти в результате трех независимых миссенс мутаций (поскольку аминокислота кодируется тремя кодонами). Поэтому мы будем рассматривать три различных варианта мутантов (табл. 13). Используя таблицу генетического кода, определите, какие изменения должны произойти в кодирующей нити ДНК у клеток дикого типа для образования каждого из мутантов. Результаты внесите в таблицу. Таблица 13 Характеристика мутаций, вызвавших замену Met на Ile в белке
Примечание. Таблица приведена для образца. Решение: В результате миссенс мутации в мутантном белке произошла замена Met на Ile. Аминокислоте Met в мРНК соответствует кодон АУГ (АТГ в кодирующей нити ДНК). Изолейцин имеет три кодона АУУ, АУЦ и АУА (АТТ, АТЦ и АТА в ДНК). Следовательно, мутантный ген имеет один из этих трех кодонов. Если Ile кодон представлен АТА, следовательно, произошла замена Г→А, что соответствует транзиции. Если Ile кодон представлен АТЦ, то произошла замена Г→Ц, что соответствует трансверсии. Если Ile кодон представлен АТТ, то произошла замена Г→Т, что также соответствует трансверсии. Ответ:
Задача 3.Имеется двухспиральная молекула ДНК, представляющая собой участок гена: 5`-ЦАЦТАТГЦТТГЦГТГГАЦГЦАТТААЦ- 3` 3`-ГТГАТАЦГААЦГЦАЦЦТГЦГТААТТГ- 5` 1). Пусть транскрипция начинается с нуклеотида, соответствующего А в мРНК, и продолжается до конца. Какова будет последовательность синтезированной мРНК? 2). Какова будет последовательность аминокислот в полипептиде после трансляции этой мРНК? 3). Как изменится структура данной молекулы ДНК, если произойдет: 3А. Таутомеризация в енольную форму первого Т на кодирующей нити в процессе репликации ДНК? 3Б. Таутомеризация в иминоформу второго А на некодирующей нити в процессе репликации ДНК? 3В. Дезаминирование третьего Ц на кодирующей нити перед репликацией ДНК? 3Г. О6- алкилирование четвертого Г на некодируюющей нити до репликации ДНК? 3Д. Делеция третьего Ц на кодирующей нити? Решение: 1. Так как верхняя нить является кодирующей, она имеет ту же полярность и смысл, что и мРНК. Следовательно, эта нить может быть использована как модель мРНК, только в мРНК вместо тиминов будут находиться урацилы. Основанию А в мРНК будет соответствовать Т в кодирующей нити, значит транскрипция начнется со второго нуклеотида и синтезированная мРНК будет иметь вид: 5`-АЦУ АУГ ЦУУ ГЦГ УГГ АЦГ ЦАУ УАА Ц- 3`. 2. Находим первый АУГ кодон, с которого может начаться трансляция и, пользуясь таблицей генетического кода, определяем аминокислотную последовательность полипептида, который будет считываться с данной мРНК: Met – Leu – Ala – Trp – Thr – His – Stop АУГ ЦУУ ГЦГ УГГ АЦГ ЦАУ УАА 3А. Таутомеризация в енольную форму первого Т на кодирующей нити приведет к тому, что он будет спариваться с Г, образуя пару Т→Г, а после второго раунда репликации образуется пара ЦГ, т.е. ТА в молекуле ДНК заменится на ЦГ. Тогда первый кодон мРНК будет не АЦУ, а АЦЦ, однако это изменение не окажет влияния на структуру полипептида, так как этот кодон при трансляции считываться не будет. 3Б. Таутомеризация в имино форму второго А на некодирующей нити приведет к транзиции А→Г;
Г Т Г А Т А Ц… Г Т Г А Т Г Ц…
После чего в мРНК эта часть будет иметь следующий вид: 5` АЦУАЦГ… Изменения в инициирующем кодоне с АУГ на АЦГ приведет к тому, что рибосомы не узнает этот кодон, и будет искать другой АУГ кодон. Полипептид синтезироваться не будет. 3В. Дезаминирование третьего Ц в кодирующей нити приведет к его превращению в У. Это приведет к транзиции Ц→Т и мРНК станет: АЦУАУГУУУ…, а полипептид будет иметь вид: Met-Phe-Ala-Trp-Thr-His. 3Г. О6- алкилирование четвертого Г на некодируюющей нити сделает похожим его на А, в результате после репликации произойдет транзиция Ц→Т в некодирующей нити, а в результате после второго акта репликации пара ЦГ заменится на ТА: 5`-Ц А Ц Т А Т Г Ц Т Т Г Ц Г… 5`-Ц А Ц Т А Т Г Ц Т Т Г Т Г… 3`-Г Т Г А Т А Ц Г АА Ц Г Ц… 3`-Г Т Г А Т А Ц Г АА Ц А Ц.., а в синтезируемом белке аминокислота Ala замениться на Val. 3Д. Делеция третьего Ц в кодирующей нити приведет к следующим изменениям: ЦАЦ ТАТ ГТТ ГЦГ ТГГ АЦГ ЦАТ ТААЦ ГТГ АТА ЦАА ЦГЦ АЦЦ ТГЦ ГТА АТТГ, тогда мРНК будет давать следующий белок: АЦУ АУГ УУГ ЦГУ ГГА ЦГЦ АУУ ААЦ Met Leu Arg Gly Arg Ile Asn Задача 4.Может ли гидроксиламин (ГА) индуцировать реверсию нонсенс мутации? Решение: Гидроксиламин вызывает мутации ГЦ→АТ или ЦГ→ТА, т.е. транзиции. В трех стоп-кодонах УАА, УГА и УАГ гуанин имеется только у двух последних, следовательно, только у них возможна замена гуанина на аденин. Однако, замена Г на А в обоих случаях вновь приведет к образованию стоп-кодона УАА. Ответ. Нет. Задача 5.Было установлено, что из 94 075 детей, зарегистрированных в клинике одного из городов Европы, обнаружено 10 случаев заболевания ахондроплазией. Двое из них имели больных родителей, а 8 детей имели здоровых родителей и, следовательно, их болезнь является результатом новой мутации. Какова частота мутирования гена ахондроплазии? Решение: Так как число детей, у которых возникли мутации – 8, и общее число гамет, участвующих в образовании зигот у всех детей равно 2´94075, следовательно, частота мутирования будет 8/2´94075 = 4,2 ´ 10-5.
Поиск по сайту: |
 5`-ЦТГ ЦЦТ ТГТ ЦАГ АЦА ТГТ АТА ЦЦЦ-3`
5`-ЦТГ ЦЦТ ТГТ ЦАГ АЦА ТГТ АТА ЦЦЦ-3`

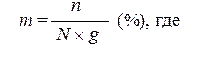
 тогда Ц А Ц Т А Т Г… Ц А Ц Т А Ц Г…
тогда Ц А Ц Т А Т Г… Ц А Ц Т А Ц Г…