|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Кроссинговер механизмі. Интерференция
Гендік мутациялар Гендік мутация - жеке гендер днқ-сының нуклеотид құрамының өзгеруi. Гендік мутация: Транзиция, Трансверсия, Инсерция, Делеция. — Транзиция– пуринді және пиримидинді нуклеотидтер орны ауысу кезінде сақталады (АТ→ ГЦ, ГЦ→АТ және т.б.) — Трансверсия – пуринді және пиримидинді нуклеотидтер ауысу кезенде орны ауысады (АТ→ЦГ, АТ→ТА және т.б.) — Инсерция – бір немесе бірнеше нуклеотид қойылуы. — Делеция– бір немесе бірнеше нуклеотидтердің жоғалуы . 3. Мейоз және оның генетикалық маңызы. Мейоз -бұл жыныс клеткаларының бөлінуі. Мейоз нәтижесінде, бір аналық біріншілік жыныс клеткаларынан (2n4c) - хромосома жиынтығы екі есеге қысқарған (1n1c) төрт жас клеткалар түзіледі. Яғни, осы бөліну арқылы гаплоидты жыныс клеткалары: жұмыртқа клеткалары мен сперматозоидтар және өсімдік споралары түзіледі. Мейоз кезеңдері: Мейоздың алғашқы бөлінуі(редукциялық) Мейоз1 Профаза1, метафаза1 анафаза1, телофаза1. Мейоздың екінші бөлінуі (эквационды) Мейоз 2 Профаза2, метафаза 2, анафаза2, телофаза2.
4. Хромосомалық мутациялар Хромосомалық мутация түрлі хромосомалық өзгерістер немесе абберацияларға байланысты болады. Бұл жағдайда хромосомалардың құрылымы өзгереді. Ондай өзгерістер хромосома ішілік және хромосома аралық болып келеді. Хромосомаішілік өзгерістерге мыналар жатады: 2.делеция — хромосоманың бір бөлігінің үзіліп түсіп қалуы; 5. Моногибридті будандастырудағы тұқым қуалау заңдылықтары Моногибридті будандастыру. Тұқым қуалаушылықты зерттегенде Мендель ең қарапайым моногибридті будандастырудан бастап, әрі қарай біртіндеп күрделендіре түскен. Мендельделдің ұсынысы бойынша гендер латын әріптерімен белгіленеді. Бір аллельді жұптың гендерін бірдей әріппен, яғни доминантты (басым) генді үлкен әріппен, рецессивті (басылыңқы) генді кіші әріппен белгілеу ұсынылған. Олай болса доминантты қасиет көрсететін асбұршақтың гүлінің қызыл түсі мен тұқымының сары түсін — А, рецессивті — гүлдің ақ түсі мен тұқымының жасыл түсін — а деп, тұқымының доминантты тегіс пішінін — В, рецессивті кедір- бұдырлы пішінін — b деп белгілейді. Ата-аналық формалар Р әріпімен белгіленеді (латынша “parents” — ата-ана). Аналықты белгісімен, аталықты белгісімен, будандастыруды Х, ұрпақтарын Ғ әрпімен (латынша “Fіlіus” — ұрпақ), гаметаларды Г, ал бірінші, екінші және үшінші ұрпақтарын Ғ1, Ғ2, Ғ3 және т.б. деп белгілейді. Сонымен, Моногибридті будандастыру деп – бір жұп (доминантты және рецессивті), қарама-қарсы белгілері бойынша ажыратылатын аталық-аналық дараларды будандастыруды айтады.
9. Аллеьді емес гендердің өзара әрекеттесуі (коопреация және комплементарлық реакциясы) Аллельді емес гендердің өзара әсерлесуі кезіндегі басты ерекшелік, ол ұрпақтарында жаңа белгінің дамуы. Комплементарлы –доминантты гендер генотипте жеке келгенде өз белгілерінің дамуын, ал гомозиготалы немесе гетерозиготалы жағдайда екі доминантты гендердің генотипте қатар келуі жаңа белгінің түзілуін қамтамасыз ететін гендерді айтады. Комплементарлы гендер екі жұп аллельді емес гендермен бақыланатын белгінің дамуын қамтамасыз етеді және доминантты гендер бірін-бірі толықтырады. Бұл типке 9:7, 9:6:1, 9:3:4, 9:3:3:1қатынастары жатады.Мысалы, тауық айдарларының пішіндерінің тұқым қуалауы. · Р: ♀ ААвв Х ♂ ааВВ (раушан пішінді) (бұршақ пішінді) (А алеллі) (В аллелі) G: Ав аВ
Р (
· Енді, белгілердің 9:6:1ажырау қатынасына мысал: асқабақ пішінінің тұқым қуалауы Р: ♀ SSdd Х ♂ ssDD (шар пішінді) (шар пішінді) (S аллелі) (D аллелі) G: Sd sD
Р (
Гаплоидия. Гаплоидия –соматикалық немесе жыныстық жасушалардың жиынтығындағы әр гомологиялық хромосомалардың жұбы тек біреуімен ғана көрініс бергенде, яғни хромосомалар санының азаюы. Соматикалық жасушаларында гомологты емес хромосомаларлың гаплоидты жиынтығы бар организмді – гаплоид д.а. Табиғи гаплоидия – споратүзетін саңырауқұлақтар, бактериялар, біржасушалы балдырлардың өмір циклінде кездеседі. 1921 жылы жоғ.сатыдағы өсімд-де ең алғашқы гаплоид табылды- сасық меңдуанада. Кейін бидай, жүгеріден табылды. Гаплоид фенотипінің ерекшеліктері: - Гаплоидтарда рецессивті гендер көрініс береді, себебі оларды доминантты гендербаспайды; - Сыртқы көрінісімен диплоидты ағазаларға ұқсас, бірақ кішірек; - Өз-өзін тозаңдандырушы гаплоидтарға қарағанда айқас тозаңданатын гаплоидтар өмір сүруге әлсіз ; - Гендер дозасының азаюына байланысты, гаплоид жасушалар көлемі ұсақ; - Мейозда толық гаметаларды түзбеген соң, гаплоидтар ұрпақ бере алмайды. Гаплоидия генетикалық мәселерді шешу үшін: ген дозасының әсерінің көрініс беруі, анеуплоидтар алу, сандық белгілердің генетикасын зерттеу үшін, геномды талдау. Гаплоидтардың гомозиготалық линияларының хромосомасын екі еселеу арқылы гибридті тұқым (мысалы, жүгері) өндірісінде өз өзін тозаңдандырған линиялармен тең келетін линияларды алу үшін және селекциялық процесті полиплоидтықтан диплоидты деңгейге ауыстыру (мысалы, картоп). Гаплоидияның ерекше формасы – андрогенез. Спермия ядросы жұмыртқажасушаның ядросының орнын баса отырып, жүгеріде стерильді аталық аналогтарын алу үшін қолданылады.
11. Аллельді емес гендердің өзара әрекеттесуі (эпистаз, полимерия, плейотропия) · Эпистаз– бір геннің ісерін екінші бір ген тежеуін, яғни көрінуіне мүмкіндік бермейтін аллеьді емес гендер. Белгінің көрінуін тежейтін гендер – эпистаздық гендер, ал әсері басылыңқы болатын гендер – гипостаздық гендер. Э.құбылысы >белгісімен белгіленеді. Егер А гені В генінің әсерін тежесе, доминантты эпистаз, ал сс гені Д генінің көрінуіне мүмкіндік бермесе, рецессивті эпистаз д.а. Домин-ы эпистаз: 12:3:1, 13:3.Рец-і эпистаз: 9:3:4. Мысалы, тауық қауырсындарының түсінің тұқымқуалауы – домин.эпистаз.Мысалы, леггорн және плимутрок тұқымды тауықтардың қауырсын-ың ақ түсі дом-ы (ССІІ) және рец-і (ссіі) ген-мен анықталады. С гені- қауырсынның боялуына, с аллелі – қауырсынның боялмауын бақылайды. І гені С генінің әсерін тежегенімен, і гені ол әсерді тежей алмайды. Р: ♀ ССІІ Х ♂ ссіі Ақ ақ G: СІ сі
Р (
Яғни, Ғ2-де алынған балапандардың ақ түсті болуы генотиптегі С генін тежеуші эпистаздық І генінің болуымен түсіндіріледі. Ал қауырсындары боялған балапандардың генотипінде С генінің әсерін тежейтін доминантты І гені болмаған. Доминантты эпистазға Ғ2-де фенотип бойынша 12:3:1 ажырау типі жатады. Мысалы, жылқыларда қара түс доминантты В генімен, ал осы геннің рецессивті в аллелі жирен түсті анықтайды. Екінші геннің доминантты С гені эпистаздық қасиет көрсетендіктен, жылық түсі сұр болады. Р: ♀ ССВВ Х ♂ ссвв Сұр жирен
Ғ1-де алынған сұр жылқыларды өзара будандастырса, Ғ2-де: 12 сұр(9/16- С-В-; 3/16- С-вв):3қара(3/16-ссВ-):1жирен(1/16-ссвв) болып ажырайды. · Кез келген бір белгінің дамуына бірігіп, әсер ететін екі немесе одан да көп аллельді емес гендерді полимерлі гендердеп, ал ондай гендердің өзара әсерлесу құбылысын полимерияд.а. Көбінесе полим.гендер сандық белгі-ің көрінуіне, яғни өлшеу,санау,талдау арқылы ғана баға беріге болатын белгілерді (өсімдіктің биіктігі, жануар. Салмағы, тауық.жұмыртқалаштығы, сүттің майлылығы, қынт қызылшасының тамыр жемісіндегі қанттың мөлшері, т.б.) бақылайды. Полим.гендер Полимерия: кумулятивті және кумулятивті емес. Кум-і емес полим. Кезінде белгінің қалыптасу деңгейі полимерлі гендердің доминантты аллельдерінің санына емес, доминантты аллеьдің генотипте болуымен анықталады. Мысалы, бидай дәнінің қызыл түсі доминантты аллеьді емес екі полимерлі Тәжірибе барысында Г.Нильсон-Эле Ғ2 популяциясындағы қызыл дәнді өсімдіктердің дәні түсінің қою қызылдан ашық қызылға дейін өзгеретіндігін байқаған. Оның жорамалы бойынша, бидай дәнінің түсінің интенсивтілігі генотиптегі доминантты аллельдерінің санына тәуелді, яғни өсімдік генотипінде доминантты аллельдер саны неғұрлым көп болса, соғұрлым белгі қоюлана түседі деп есептеді:
Кумул-і полим. – белгінің көріну деңгейі ағза генотипіндегі полимерлі гендердің домин-ы аллельдерінің санына тәуелді болады. Мысалы, адам терісінің түсі екі жұп полимерлі доминантты Ғ2-де тері түсінің ажырауы фенотип бойынша 1:4:6:4:1қатынасында болады. · Плейотропия –бұл әртүрлі мүшелер мен ұлпаларға бір геннің тәуелсіз немесе автономды әсері және бір геннің бірнеше белгілердің түзілуіне әсері. Мысалы, дрозофиладағы көздің «ақ» түсін анықтайтын ген бір мезгілде, ішкі мүшелердің түсінің өзгеруі мен тіршілік қабілетінің төмендеуіне әкеледі. Модификаторлы гендер – ағза генотипіндегі кейбір гендердің нақты белгінің дамуына емес, басқа бір гендердің даму жағдайларые өзгерте алу қасиетін айтады. Мұны кезкелген белгіні немесе қасиетті анықтайтын генотиптегі негізгі гендердің әртүрлә жағдайда көрінуіне модификаторлы гендердің әсерімен түсіндіріледі. Гендердің нақты белгі немесе қасиет түрінде жүзеге асуы, сонымен қатар олардың фенотиптегі көріну деңгейлері белгілі деңгейде нақты генотип пен сыртқы орта жағдайларына тәуелді болады. Геннің фенотиптегі көріну мүмкіндігін Н.В.Тимофеев-Ресовский пенетранттылықдеп, ал геннің белгіге әсер ету деңгейін- экспрессивтілікдеп атауды ұсынды. Пенетранттылық даралардағы геннің белгі немесе қасиет түрінде көрінуі пайызбен, ал экспрессивтілік нақты геннің әсерінен көрінетін белгінің орташа мәнімен өлшенеді.
19.Ақуыз биосинтезі Ақуыз биосинтезі тізбегінде жазылған ақпараттың колинеарлы полипептид тіршіліктің ең маңызды қасиеттерінің бірі, себебі тіршілік процесіңде ақуыз молекулалары әр түрлі биохимиялық реакцияларға түсіп ыдырап жойылып отырады, ал олардың орнын толтыру тек жаңа ақуыз молекулаларының синтезделуі нәтижесінде жүзеге асады. А-РНҚ молекуласының нуклеотидтер молекуласының аминқышқылдары ретіне берілуін, яғни ақуыз синтезін, трансляция деп атаймыз.Трансляция процесі полипептидтің N ұшынан басталып С ұшына қарай жүреді және ол метионин амин қышқылынан басталады. Яғни, кез келген а-РНҚ молекуласының бас жағында метионин амин қышқылына сәйкес келетін (колинеарлы) код-инициаторлық код болады (АУГ). Полипептид синтезінің инициялануы ерекше т-РНҚ-инициаторлық т-РНҚ-ның қатынасуымен жүзеге асады. Бұл процеске 3 инициациялаушы факторлар — ІҒр ІҒ2, ІҒ3 қатынасады. Ақуыз биосинтезінің жалғасуы, немесе элонгация сатысы, 2 фактордың қатысуымен жүреді — ЕҒ-1, ЕҒ-2, ал белок синтезінің терминациялануы кодон-терминаторларға және терминациялық факторларға — КҒ-1, КҒ-2-ге байланысты. Кодон терминаторлардың 3 түрі белгілі УАА, УАГ, УГА.Трансляция процесіне 3 РНҚ — үшеуі де қатынасады. А-РНҚ-ақуыз синтезі үшін матрица (қалып) қызметін атқарса, рибосомалық РНҚ рибосомалардың құрамына кіреді. Рибосомалар цитоплазманың ең ұсақ субмикроскопиялық органелла-лары болып табылады. Ақуыз синтезі жүруі үшін рибосомалар а-РНҚ-мен байланысады. Рибосома 2 бөлшектен тұрады: рибосоманың кіші бөлшегі — 308, рибосоманың үлкен бөлшегі — 508. Әдетте, ақуыз синтезі жүрмеген кезде, рибосоманың екі бөлшегі бір-бірінен бөлек кездеседі, олардың бір-бірімен қосылып біртүтас органоид — рибосома түзілуі ақуыз биосинтезі басталған кезде жүзеге асады. А-РНҚ алғаш рибосоманың кіші бөлшегімен қосылады, содан кейін оларға иницияция факторлары ІҒ1, ІҒ2, ІҒ3 және т-РНҚ — аминоацилметионин қосылып инициа-торлық кешен түзіледі.Инициаторлық кешен түзілгеннен кейін оған рибосоманың үлкен бөлшегі келіп қосылады. Рибосоманың үлкен белшегінде 2 учаске болады: пептидиддік бөлік — П бөлік, аминоацилдік болік — А бөлік. Рибосоманың үлкен бөлшегінің П бөлігіне формилденген метионинмен инициаторлық т-РНҚ орналасады, ал А учаскеге а-РНҚ-ның екінші кодонына сәйкес келетін антикодоны бар т-РНҚ өзінің амин қышқылымен орналасады.Трансляция процесінде а-РНҚ кодондары амин қышқылдарымен тікелей әрекеттесе алмайды. Олардың арасындағы байланыс тасымалдаушы РНҚ (т-РНҚ) арқылы жүзеге асады. Әрбір амин қышқылының тек өзіне ғана сәйкес т-РНҚ-сы болады, сонымен кем дегенде т-РНҚ белгілі — 20-амин қышқылының 20 т-РНҚ-асы және инициаторлық Т-РНҚ. Т-РНҚ-лар — т-РНҚфен, т-РНҚАлан, т-РНҚСеР, т-РНҚИзол, т-РНҚАсп т.с.с деп аталынады. Барлық прокариоттардың және эукариоттардың т-РНҚ молекулалары 80-ге жуық нуклеотидтерден қүралған және олар-дың соңғы құрылысы бір-біріне ұқсас болып келеді. Олардың молекулаларының соңғы құрылысы беденің 3 құлақты жапырағына ұқсас. Т-РНҚ молекуласының 2 ерекше қасиеті белгілі: 1) т-РНҚ-ның 3′-ұшында ерекше рЦ рЦ рА-ОН-тан түратын бос нуклеотид тізбегі болады, осы ұшымен т-РНҚ-ға сәйкес келетін амин қышқылы ковалентті байланысады; 2) т-РНҚ-ның қарама-қарсы ұшында 3′ нуклеотидтен тұратын антикодон болады.Т-РНҚ-ның негізгі қызметі — өзіне сөйкес амин қышқылын «танып» 3′-ОН ұшына қондыру жөне а-РНҚ-ның антикодонға комплиментарлы кодонын «танып» өзі алып келген амин қыш-қылын полипептид тізбегіне орналастыру болып саналады Амин-қышқылының өзіне сәйкес т-РНҚ молекуласының 3’—ОН үшына ковалентті байланысуы ерекше аминоацил т-РНҚ-синтетаза ферментінің қатысуы арқылы жүреді. 20 амин қышқылына сәйкес 20 аминоацил т-РНҚ синтетаза ферменттері болады. Аминоацил т-РНҚ синтетаза ферменттері ездеріне сәйкес келетін амин қышқылдарымен т-РНҚ-ның бір-бірін «танып», амин қышқылының т-РНҚ молекуласының З1-ОН ұшына коваленттік байланысуын қамтамасыз етеді. Бұл про-цесс 2 сатыдан тұрады. Алғашқы сатыда амин қышқыддары ак-тивтенеді: Екінші сатыда активтенген амин қышқылы-өзіне сәйкес т-РНҚ-ның ЗЮН ұшымен байланысады. Аминоацил-т-РНҚ-синтетаза ферменттері трансляция процесінің қателіксіз жүруін, яғни а-РНҚ кодына колинеарлы (сәйкес) полипептид молеқуласының синтезделуін, қамтамасыз етеді. Демек, трансляцияның қателіксіз жүруі аминоацил-т-РНҚ-синтетаза ферменттерінің өздеріне сәйкес амин қышқыл-дарын «танып» оларды тиісті т-РНҚ-ға қондыруына байланыс-ты болады. Сонымен қатар, аминоацил-т-РНҚ — синтетаза ферменттері бақылаушылық қызметін де атқарады. Мысалы, изо-лейцил-т-РНҚ — синтетаза ферменті өзіне сәйкес т-РНҚ ор-нына басқа — «жат» т-РНҚ, айталық т-РНҚФен, «таныды» делік. Осының нәтижесінде изолейцин-т-РНҚИзол орнына изолейцил-т-РНҚфен кешені пайда болар еді. Осы қателіктің нәтижесінде синтезделуші ақуыз молекуласында фенилаланин амин қышқылының орнына изолейцин амин қышқылы орналасар еді. Бірақ, фенилаланин-т-РНҚ-синтетаза ферменті өзінің т-РНҚФЕН-сымен жат амин қышқылының қосылғанын сезіп қалып қате кешенді — изолейцин амин қышқылына және т-РНҚфен-ге ыдыратады. Аминоацил-т-РНҚ-синтетаза ферменттерінің екінші бақы-лаушылық қызметі амин қышқылдардың активтену сатысында байқалады. Айталық, изолейцил-т-РНҚ-синтетаза ферменті изо-лейцин амин қышқылының орнына валин амин қышқылын ак-тивтендірді делік.Изолейцил-т-РНҚ-синтетаза+валин+АТФ—>(валилацил—АМФ) изолейцил-т-РНҚ-синтетаза+Ррі. Сонда, келесі сатыда «қате» түзілген кешен т-РНҚИзол мен әрекеттескенде валил-т-РНҚИзол пайда болуының орнына валилацил-АМФ, еркін валин амин қышқылына және АМФ-ге гид-ролизденеді. (Валилацил-АМФ)изолейцил-т-РНҚ-синтетаза+т-РНҚИзол->валин,АМФ+т-РНҚИзол+изолейцил-т-РНҚ-синтетаза.Сонымен, аминоацил-т-РНҚ-синтетаза ферменттері нақтылы амин қышқылдарын өздеріне тиісті антикодондармен байланыстырып генетикалық ақпараттың трансляциялану процесінде маңызды рөл атқарады. Сонымен қатар, бұл ферменттердің қос қабат бақылаушылық қызметі нәтижесінде трансляция процесінің қателіксіз жүру деңгейі едәуір жоғарылайдыАминоацил -т-РНҚ кешені пайда болғаннан кейін олар рет-реттерімен тізіліп орналасып, амин қышқылдар арасында пептидтік байланыс пайда болады. Оны рибосоманың үлкен бөлшегіңце кездесетін ерекше фермент — пептидил — трансфераза ферменті қалыптастырады. Екі амин қышқылдары арасында пептидтік байланыс пайда болғаннан кейін рибосома бір кодонға солға қарай жылжиды, осының нәтижесінде инициаторлық кешен рибосомадан сыртқа шығады, А учаскедегі кешен П учаскеге өтеді, ал А учаскесіне а-РНҚ-ның үшінші кодонына сәйкес келетін т-РНҚ өзінің амин қышқылымен келіп орналасады, со-дан кейін екінші және үшінші амин қышқылдары арасыңца пептидтік байланыс пайда болады. Бұл жерде элонгация факторлары — ЕҒ-1, ЕҒ-2 қатынасады. Содан кейін рибосома тағы бір кодонға солға жылжып А учаске босайды. Осылайша ақуыз син-тезі А учаскеге а-РНҚ-ның терминаторлық кодондары келіп жеткенге дейін жалғаса береді. Терминаторлық кодондарды (УАА, УАГ, УГА) арнайы терминация факторлары — КҒ-1, КҒ-2 «таниды». Терминаторлық кодоңцарға сәйкес келетін амин қышқылдары болмағандықтан А — учаске бос қалып, пептидил-трансфераза ферменті пептидтік байланыс жасай алмайды, сондықтан рибосома 2 бөлшекке ыдырайды, синтезделген полипептид бөлініп шығады. Осылайша ақуыз синтезі аяқталады. 22.Жыныспен тіркес белгілердің тұқым қуалауы Гендері жыныс хромосомаларда орналасқан белгілерді жыныспен тіркескен белгілер деп атайды. Ал белгілердід жыныстық хромосомалары (X және Ү) арқылы ұрпақтан-ұрпаққа берілуін жыныспен тіркескен белгілердің тұқым қуалауы деп атайды. Бұл құбылысты Т.Морган дрозофила шыбынына тәжірибе жүргізгенде ашкан.Дрозофила шыбынының көзінің қою қызыл түсті белгісі (W), ақ белгісінен басым (w). Гомозиготалы қызыл түсті аналық шыбынды (XWXW), ақ көзді аталық шыбынмен (XWY) шағылыстырғанда, Fх-де ұрпақ шыбындардың көздерінің түсі қызыл болған. Ғ2— ұрпақта белгілер 3 : 1 қатынасында ажыраудың орнына, барлық аналық шыбындар қызыл көзді, аталық шыбындардың жартысы қызыл, қалған жартысы ақ көзді болған. Ал ақ көзді аналық шыбынды, көзінің бояуы қою қызыл түсті аталығымен шағылыстырғанда, FL — ұрпақта аналық шыбындар әкесінен қызыл түсті, аталық. Шыбындар анасынан ақ түсті алатындықтан, белгілер айқас тұқым куалайды. Ғ2— ұрпақта жыныстык хромосомалардың тұқым қуалауына сәйкес, аналық шыбындардың жартысы ақ көзді, жартысы кызыл көзді, осымен қатар аталық шыбындардың да жартысы қызыл, жартысы ак көзді болады.Жыныстық хромосомаларда орналаскан гендер аныктайтын белгілердің тұқым қуалауы, Мендель анықтаған белгілердің ажырауынан өзгеше болады. X және Ү хромосомалардың гомологті емес бөліктері бар, сондықтан да X хромосомадағы гендердің аллельдері Ү хромосомада болмайды. Керісінше, Ү хромосомада бар гендердің аллельдері X хромосомада жоқ. Гендердің мұндай күйін гомизиготалы деп атайды.Егер ген X хромосомада орналасқан болса, онда ген әкесінен қыздарына, шешесінен қыздарына және ұлдарына белгілер теңдей беріледі. Белгілердің шешесінен ұлдарына, ал әкесінен кыздарына берілуі Крисс-кросс тұқым қуалау деп аталады.Белгілерді бақылайтын гендер Ү хромосомада орналасса, онда X хромосомада оған сәйкес бөлік болмағандықтан, әкеден тек ұлдарына беріледі. Себебі қыздарына хромосома берілмейді. Мысал ретінде адамда аяқ саусақтарының арасында жарғактық болуы, құлақтың ішінде шоқ жүннің болуы, тек әкеден ұлдарына берілетін белгілер. Құстарда, кейбір балықтардың түрлерінде және көбелектерде көптеген белгілер жыныспен тіркесіп берілетіні анықталған. Бұл жағдайда белгілерді анықтайтын гендер X хромосомада орналасқан. Бірақ аналық гетерогаметалы да, аталық гомогаметалы болады.
26. Организмнің жеке дамуының генетикалық бағдарламасы. Онтогонез(грек. on – табыс септігінің жалғауы, ontos – нағыз, нақты және генез) – организмнің жеке дара дамуы. Онтогенез ұрық болып түзілуінен бастап, тіршілігінің соңына дейінгі барлық өзгерістердің жиынтығы. Онтогенез терминін неміс биологы Э.Геккель үсынған (1866). Онтогенез барысында дамып келе жатқан организмнің жеке мүшелері өсіп, жіктеледі және бірігеді. Осы күнгі көзқарастар бойынша Онтогенезге бастау болатын жасушаның ішінде организмнің одан әрі дамуын анықтайтын белгілі бір тұқым қуалаушылық бағдарламасы – код түріндегі мағлұмат сақталады. Бұл бағдарлама бойынша Онтогенез барысында ұрықтың әрбір жасушасындағы ядро мен цитоплазманың әсерлесуі; сондай-ақ, ұрықтың әр түрлі жасушалары мен жасуша кешендерінің өзара әрекеттесулері жүзеге асады. Тұқым қуалау аппараты өзіндік белок молекулаларының синтезделуін кодтау (белгілеу) арқылы морфогенетикалық процестердің жалпы бағытын ғана анықтайды, ал олардың нақтылы жүзеге асырылуы белгілі дәрежеде (тұқым қуалаушылық нормасы шеңберінде) сыртқы факторлардың әсеріне тәуелді болады. Организмдердің әр түрлі топтарында Онтогенездің тұқым қуалаушылық бағдарламасының мүлтіксіз орындалу деңгейі мен оның реттелу шегінің мүмкіншілігі мол. Онтогенез — бір тірі ағзаның ұрық біте бастағаннан өлгенге дейінгі даму үдерісі. Онтогенез екіге бөлінеді: эмбриогенез (тууға дейінгі тіршілік) жөне постэмбриогенез (туу соңынан кейінгі тіршілік).Органогенез — мүшелердің түзілу үдерісі. Адамның, барлық омыртқалы жануарлардың жөне кейбір омыртқасыз жәндіктердің ұрығы реттеліп дамиды. Яғни ұрық денесінің қай бөлігі қай жасушалардан дамитыны кештеу сатыларда (гаструлаларда) анықталады және қоршаған ұрықтық жасушалардың өзара өрекеттесуіне төуелді болады. Бұдан езге ұрықтың теңбілді даму типінің жолы бар, мұнда дененің қай бөлігі қандай жасушадан дамитыны 2-немесе 4-ұрықтық жасушалар сатыларында анықталып та қойған. Ф.Мюллер және Э.Геккель 1864 жылы биогенетикалъщ заңды: «Онтогенез қысқа жөне тез қайталаған филогенез» деп тұжырымдады. Ұрық өз түрінің эволюциясын қысқа қайталағандай болып, жүйелік топтардың ататектік ұрықтық сатыларын қайталайды деген дөлелдемелер өте көп 27. Жыныс айырудың баланстық теориясы. 1922 жылы К.Бриджес дрозофиланың үшеселенген хромосомалар жиынтығы бар үшплойдты 3Х+3А ұрғашыларын тапты. Кейбір мұндай ұрғашы шыбындар кәдімгідей өсімтал болып шықты. Оларды диплоидты ХУ+2А еркек шыбындармен будандастырғанда ұрпақтарының ішінде жынысаралық белгіердің – интерсекстердің пайда болатыны байқалады. Морфологиялық, цитологиялы және генетикалық зерттеулер мұндай ұрпақтардың әртүрлі қатынастағы аутосомалар мен жыныстық хромосомалары бар 8 түрін айқындады. 1. 3Х:3А 2. 2Х:2А 3. 2Х+У:2А 4. 2Х:3А 5. 2Х+V:3А 6. ХУ:2А 7. 3Х:2А 8. ХУ:3А Әртүрлі хромосомалар жиынтығы бар шыбындардың шығу себебі, үшплоидты ұрғашы дрозофилалардың хромосомаларының мейозда дұрыс ажыраспауынан болған. Әртүрлі хромосомалар балансы бар шыбындар: 1. Қалыпты еркек және ұрғашы 2. Аралық белгілері бар – интерсекстер 3. Шамадан тыс дамыған белгілері бар еркек шыбындар (сверхсамец) 4. Шамадан тыс белгілері дамыған ұрғашы ұрпақтар (сверхсамка)берді Сөйтіп тұқымның жынысы Х-хромосома мен аутосомалар жиынтығының балансына байланысты анықталатыны белгілі болды. Мысалы, (2Х:3А) аутосомалар құрамы көп болса еркек интерсекс шыбындар шығады. Тең балансты диплоидты немесе үшплоидты (2Х:2А немесе 3Х:3А) болса ұрғашы болып шығады. Х- хромосома санының азаюы (2Х:2А) еркек шыбындардың шығуына мүмкіншілік береді. Егер аутосомалар жиынтығы үшке дейін өзгеріп, Х-хромосома біреуі ғана болса(Х:3А), онда «шамадан тыс еркек» - еркектің белгілері шамадан тыс дамыған , бірақ ұрықсыз организмдер пайда болады. Керісінше Х-хромосомалар көбейіп, аутосомалар диплоидты күйде қалса (3Х:2А) «шамадан тыс ұрғашылар» - жұмыртқа клеткалары және басқада жыныстық белгілері өзгерген ұрғашылар пайда болады. Осы зерттеулердің нәтижесінде К.Бриджес шыбындардың жынысы екі Х немесе ХУ хромосомаларға байланысты емес, олардың жыныстық хромосомалар мен аутосомаларының сандарының қатынасына байланысты деген қорытындыға келді. Ұрғашы жыныстың негізгі даму бағыты Х- хромосомада орналасқан, ал еркек жыныстылардікі – аутосомаларда. Бұл мынадан көрінеді: егер хромосомалар балансы(немесе жыныс индексі) Х:А =1 болса ұрғашы, Х:2А =0,5 еркек болады. Хромосомалар балансының 1-ден 0,5 ке дейінгі қатынасы аралық жыныс, яғни интерсекстерді анықтайды.Үш Х-хромосаманың екі аутосомаға қатынасы 3Х:2А =1,5 шамадан тыс ұрғашылар , керісінше аутосомалар бір Х- хромосоманың үлесіне көбейгенде Х+У:3А=0,33 шамадан тыс еркек шыбындар пайда болады, бұл формаларды «суперсекстер» деп атайды. Сөйтіп, К.Бриджес жыныстық теорияны тұжырымдады: жыныстық белгілердің дамуы оған жауапты гендердің балансына байланысты. 28.Геномдар бірегейлігі және гендердің дифференциальді экспрессиясы Геном(ағылшынша genome, грекше genos — шығу, тек) — хромосомалардың гаплоидты (сыңар) жиынтығында шоғырланған гендердің бірлестігі. Геном терминін 1920 жылы неміс биологы Г. Винклер енгізді. Гаплоидты жиынтық көбінесе жыныс жасушаларына тән, ал сомалық (дене) жасушаларында хромосомалардың диплоидты (екі еселенген) жиынтығы болады. Кейде хромосомалардың саны қалыпты диплоидты жағдайдан артып кетеді. Егер гаплоидты жиынтықтан Геном үш не төрт есе артық болса, триплоидты және тетраплоидты, ал бір Геном ағзада бірнеше рет қайталанса, автополиплоидты, ал әр түрлі біріккен ағза аллополиплоидты деп аталады. Хромосомалардың жиынтығы еселеніп, артқан сайын Геном саны да өсіп отырады. Әдетте диплоидты клеткада хромосомалар жұп болып келеді. Себебі, ұрықтану кезінде оның бір сыңары аналық гаметадан, екіншісі — аталық гаметадан беріледі, яғни бұл Геномдар сәйкес (гомологты) болады. Сөйтіп екі гаплоидты жасушадан бір диплоидты жасуша түзіліп, жаңа ағза қалыптасады. Әр хромосомада тізбектеліп орналасқан гендердің өзара дәл келуін екі Геномның сәйкестігі деп атайды. Туыстығы қашық буындарда барлық немесе бірнеше Геномдар арасында сәйкестік болмайды. Бұл тұрақтылық бұзылып, белгілі бір факторлардың әсерінен хромосома санының өзгеруін (мысалы, артып, не кеміп кетсе) геномдық мутация деп атайды.Тірі ағзаларда хромосомалардың саны тұрақты болады. Мысалы адамда — 46, маймылда — 48, қиярда — 14, жүгеріде — 20, қатты бидайда — 28, жұмсақ бидайда — 42, дрозофила шыбындарында — 8, т.б. Организм эволюциялық дамуында неғұрлым жоғары сатыда тұрса, соғұрлым олардың Г-ында ДНҚ көбірек болады. Дифференциация ‐дамып келе жатқан организмнің құрылыстық, биохимиялық өзгерісі. Оның нəтижесінде клетка, ұлпа, мүше жəне организм бір түрліден көп түрліге айналады. Сонымен дифференциация – жаңа ферменттік жүйенің, ерекше белоктардың, мембраналардың жаңа қасиеттерінің түзілуі жəне физикалық, химиялық өзгерістері, жаңа ішкі ортаның түзілуі. Клетка дифференциациясына тән ерекшелігі ол қайтымсыз сол немесе соған ұқсас клеткалардың типтеріне алып баруымен сипатталады. Бұл процестің детерминация деген атауы бар және генетикалық қадағалануда ,тирозинкиназа рецепторы арқылы пептидтік өсімділік факторларымен іске асатын сигналдардың негізінде клеткалардың дифференциация және детерминациясымен әсерлесіп , реттеледі. Қазіргі таңдағы цитодифференцировканың негізгі механизмі болып дифференциальді гендердің экспресиясы саналады. Гендердің дифференциальді экспресиясының реттелуінің деңгейі ген - полипептид – белгі бағытындағы информацияның реализация этаптарына байланысты және клеткаішілік процестерді ғана емес, сонымен қатар ұлпалық және ағзалық процестерді де қамтиды. Гендердің дифференциациясы әр түрлі уақытта әсер етеді деп есептеледі . Бұл мРНК сы әр түрлі және әр түрлі дифференциацияланған клеткалардың транскрипциасындағы гендердің репрессиясы және дерепрессисында орын алады . Мысалы , теңіз кірпісінің бластоцистасының РНК-ғы транскипсирленетін гендердің саны – 10% ға тең , ал атжалмандардың бауырының клеткаларындағы саны да – 10 % ға тең , ірі мүйізді жануарлардың тимусының клеткаларындағы саны – 15 % ға тең .
29-сұрақ.Жыныстардың ара-қатынасы. 1:1 арақатынасын қамтамасыз ететін жынысты анықтаудың генетикалық механизмі болады. Жыныстардың генетикалық анықталған арақатынасын алғашқы арақатынас деп атайды. Алайда даму процесінде аталық және аналық гаметаларының тіршілік қабілеттілігі біркелкі болуына байланысты және басқа себептердің салдарынан, жыныстардың арақатынасы өзгеруі мүмкін. Жеке даму процесінде әр түрлі факторлардың әсерінен болған арақатынас өзгерісін жыныстың қосымша ара-қатынасы деп атайды. Жыныстың қосымша арақатынасы көбінесе аналықтардың басым болуы жағына қарай өзгереді, мұны аталық жыныс өкілінің тіршілік қабілеттілігінің нашарлығымен түсіндіруге болады. Осы жынысты реттеу проблемасы мал шаруашылығының өнімін артық беретін жыныстың бір түрін алуға негізделген. Мысалы, сүт шаруашылығында тек ұрғашы малдың, ет беретін еркек малдың туғаны ұтымды. Себебі олар жылдам өседі және етті көп береді. Сүтқоректілердің жыныстық арақатынасын реттеу ,олардың сперматозоидтарын екі фракцияға: біріншісі – Х-хромосомасы барлар, екіншісі – У-хромосомасы барлар деп бөлу арқылы жүргізілуі тиіс. Аналықтарда осы фракциялардың бірумен ұрықтандыру арқылы, бір жынысты ұрпақ алуға болады. Партеногенез – организмнің ұрықтандырусыз дамуы. Ұрпақтардың бір ғана жынысын алу эмбриондар ұрығының аталық(андрогенез) немесе аналық гаметаларынан(гиногенез) дамуына байланысты. 30-сұрақ. Популяция және оның генетикалық құрылымы. Популяция дегеніміз – өмір сүретін ортақ мекені бар, сол жердің жағдайына бейімделген, туыстас жиынтықтықтардан оқшауланған және өзара шағылыса алатын бір түрге енетін организмдердің жиынтығын айтады. Популяция түрдің бір бөлігі. Басқа популяцияларды салыстырса, сол популяциядағы организмдердің фенотипі, генотипі, өмір сүргіштігі әртүрлі болып келеді. Әдетте популяция тұйық топ. Оған басқа жердек әкелмейді, оның ішінен әкетілмейді, сондықтан популяцияда жұп таңдау өз ішінде жүреді. Әрбір популяция өзінің генофондымен , яғни құрамына кіретін ген аллельдерінің жиынтығымен сипатталды. Популяциялар эфолюцияның қозғаушы үш факторы: тұқым қуалаушылық, өзгергіштік және сұрыптаудың өзара әрекеттесуі негізінде тіршілік жағдайларының әсерімен қалыптасады. Популяциямен қоса генетикада таза линия деген ұғым бар. Таза линия – бір атадан тараған және оған генотипі жағынан өте ұқсас ұрпақтар. Негізінде таза линия малда жоқ, ол өз бетімен тозаңданатын бір өсімдіктің ұрпақтары. Жеке организм өздігінен тозаңданғанда жаңа нәсілдің, сорттың, тұқымның басы бола алады. Популяциялардың қалыптасу процестері мен олардың өскелең дамуы микроэволюция құрайды. 31-сұрақ. Жыныспен тіркес белгілердің тұқым қууы. Организмнің белгілері мен қасиеттерінің жиынтығы – жыныс арқылы ұрпақ таралып, тұқым ққуы арқылы нәсілдік хабар ұрпаққа беріледі. Организмнің әрбәр қасиеті сияқты жыныс та тұқым қуу арқылы анықталған. Жыныс айрмашылығының шығу тегі жыныстық көбеюмен тығыз байланысты. Жыныс – жыныстық жолмен көбеюіне байлансыты организмнің морфологиялық, физиологиялық ерекшеліктері. Жыныстың жіктелуі ген қасиетіне байланысты. Жыныс детерминациясын негізгі үш түрге бөледі: 1)эпигамдық – жыныстық организмнің өсіп даму(онтогенез)процесінде жіктелуі. Бұл жағдайда жыныстың анықталуы көбінесе қоршаған ортаға тәуелді. 2)Прогамдық – жыныс жіктелуі ата-аналардың гаметогенезі кезінде анықталады. 3)Сингамдық – болашақ ұрпақтың жынысы гаметалар қосылған кезде анықталады. Бұл жыныс детерминациясының ең көп тараған түрі болғандықтан, жануарлардың барлығына тән қасиет. Жыныс хромосомалары Х және У хромосома түрінде бөлінеді. Әйел – ХХ, ер адам – ХУ. Жыныстық хромосомалары бірдей гаметалардан түзілсе оны гомогаметалы жыныс деп атайды. Еркекте сперматозоидтардың екі түрі – Х немесе У-хромосомасы мейоз механизміне сәйкес тең шамада түзіледі., сондықтан дрозофиланың аталық жынысын гетерогаметалы деп атайды. Гетерогаметалы аналық жыныстар да кездеседі: аналығы zw, аталығы zz. У хромосомасы болмайтын жануарлар да болады, және аталығының жыныс жалғыз ғана Х-хромосомасымен (ХО) айқындалады.
32.Аутбридинг және имбридинг Селекцияда будандастырудың әртүрлі жүйесі қолданылады.Ол туыстық будандастыру-ибридинг, және туыстық емес-аутбридинг.Туыстық будандастыру генотиптері жағынан бір-біріне жақын организмдердің арасында болатындақтан олардан көбінесе гомозиготалы ұрпақ алынады.Мұндай будандастыруды ұзақ уақыт пайдаланып , жануарлар мен өсімдіктердің бойында пайдалы қасиеттері бар гомозиготалы таза линиаларды алуға болады. Попляция құрамындағы рецессивті гендердің көпшілігі организм үшін тиімді бола бермейді, сол себепті егер туыстық будандастыру жиі қайталанса , сорт немесе тұқым ағындайды. Сондықтан туыстық емес будандастыру әдісі қолданылады. Мұндай жағдайда әртүрлі линияларға жататын дарақтар будандастырылады, соның нәтижесінде олардың ұрпағының тұқым қуалайтын қасиеттері жақсарады. 33.Жыныстық хромасоманың ажырамауы К.Бриджес 20жылдардың бас кезінде дрозофила жыныстық белгісінің дамуы Х хромосома мен аутосомалардың ара қатынасына байланысты өзгертетіндігін байқады.Бұл шыбында кейде триплоидтық хромосома жиынтығы бар аналықтар пайда болады.Кейбір триплоидты аналықтар ұрпақ береді, бәрақ меоз кезінде олардың хромасомалары дұрыс ажырамайды.Мұндай триплоидты аналықты қалыпты диплоидты аталықпен будандастырғанда жыныстық хромасомалар мен аутосомаларының ара қатынасы әр түрлі сегіз типті дарақтар алынған. Олардың арасында қалыпты аналық және аталықтармен қатар гипертрофты особьтар да , сол сияқты асыра аналық және асыра аталықтар да пайда болған .Осы жүргізілген зерттеулердің негізінде Бриджес дрозофилада аналық жыныс екі Х-хромосомамен , ал аталық жыныстың дамуы ХУ-хромосомалармен анықталмайды, олар Х-хромосома санымен аутосомалар жиынтығын ара қатынасына немесе жыныстық индекске байланысты деген қорытындыға келді. Кейде ата-аналардың жыныс клеткаларының түзілуі кезінде жыныстық хромосомалар ажырамайды, соған байланысты адамда күрделі психикалық аурулар пайда болады,Мейоз кезінде жыныстық хромосомалардың ажырамауы себепті жұмыртқа клеткасында жалғыз Х-хромасоманың орнына екеу болады немесе бәреуі де болмайды. Мысалы: Шершевский Тернер синдромы-әйелдерде болатын ауру.Жыныс бездері болмайды,екінші жыныстық белгілер толық жетілмеген ,ұрпақ бермейді. Клайнфельтер синдромы-ер адамда болады. Жыныс бездері жетілмейді, қол-аяқтары денесіне сәйкес дамымаған. 34.Клетканың бөліну түрлері Митоз бен мейоз Клеткалық циклдің үш басты кезеңі белгілі: 1. Интерфаза. Бұл кезең жылдам синтез және өсуден тұрады. Интерфаза соңында ДНК репликациясы және екіеселену жүреді. 2. Митоз /кариокенез/. Бұл ядроның бөліну процесі және хроматидтер бір-бірімен ажырап жаңа клеткаға тепе-тең бөлінеді. 3. Цитокенез – цитоплазманың екі жаңа клетка арасында бөлінуі. Интерфаза 3 фазаға бөлінеді: G1, S, G2. G1- фазасы жылдам синтезбен сипатталады: митохондрия, Гольджи комплексі, ЭПТ, ядрошық, рибосомалардың түзілуі. Сонымен қатар клеткада құрылымдық және функциональды белоктар түзіліп клетканың өсуі жүреді. S фазада ДНК репликациясы немесе екіеселену процестері жүреді. Гистонды белоктардың синтезі жүреді. Әрбір хромосомалар екі хроматидтеге бөлініп өзара центромера немесе кинетохормен байланысады. G2 фазада биосинтез процесі жүреді және АТФ немесе макроэргтар түрінде энергия сақталады, митохондрия бөлінеді. Бөліну ұршығы түзіледі және центриолдер репликациясы жүреді. Клетканың М фазаға өтуі М қолдаушы факторлармен бақыланады. Интерфаза біткеннен кейін кариокенез немесе митоз басталады. Митоз бес кезеңге бөлінеді. Митоздың кезеңдері: профаза, прометафаза, метафаза, анафаза, телофаза. Прометафаза. Бұл фазада мембрана қабыршағы еріп майда мембрана көпіршігіне айналады. Бөліну ұршығының микротүтікшелері ядро аймағына еніп кинетохорамен бекінеді. Бөліну ұршығының микротүтікшелері үш категорияға бөлінеді – кинетохорлы, полюсті, астральді. Микротүтікшелер хроматидтердің екі жағына бекініп хромосомаларды жылдам қозғалысқа алып келеді. Микротүтікшелер саны әрбір кинетохорамен байланысқан, адам баласында 20-40 тең болады. Метафаза. Микротүтікшелер хромосомаларды экватор аймағына алып келеді, метафазалы пластинка түзеді және клетка экваторына перпендикуляр орналасады. Анафаза. Бұл өте қысқа фаза. Әрбір центромера ыдырап екіге бөлінеді және бөліну ұршығының жіптері центромераны қарама-қарсы полюстерге ажыратады. Ажырау сигналы Са ионының жылдам босануына байланысты болады. Бір-бірінен бөлінген хроматидтерді центромера өзімен бірге тартып хромосомалар түзіледі. Хромосомалардың ажырау процесіне кинетохорлы микротүтікшелер /қысқару/, полюсті /ұзару/, астральды /полюсті итеруші/ қызмет атқарады. Телофаза. Хромосомалар клетка полюстеріне келіп, ширатылуы тарқатылады. Бөліну ұршығы жойылады, центриоли репликациясы жүреді. Әрбір хромосомалар айналасында ядро қабықшасы түзіледі. Ядрошық түзіліп цитоплазманың бөлінуі цитокенез құбылысы өткеннен кейін хромосома жиынтығы диплоидты екі клетка түзіледі. Мейоз- бұл жыныс жасушасының бөлінуі және диплоидты хромосома саны гаплоидтыға төмендейді. Мейоздың жалпы үлгісі: жасушада бір рет хромосоманың екі еселенуі /ДНҚ-репликация/, бөліну 2-і циклдан тұрады: 1-ші және 2-ші мейозды бөліну нәтижесінде бір диплоидты жасушадан төрт гаплоидты жасуша түзіледі. Жасушаның бөліну фазалары: интерфаза, профаза, метафаза, анафаза, телофаза және бөлінуден кейін тағы да интерфаза. · Кроссинговер – гомологиялық хромосомалар арасында бөліктердің алмасуы, бірінші мейозды бөлінудің профазасында жүреді (профаза I), оның келесі этаптары бар:лептотена, зиготена, пахитена, диплотена, диакинез
Лептотена фазасында хромосома конденсациясы жүреді, олар ақуызды жіп және ұзын жұқа талшық түзеді. Әрбір хромосома екі соңымен ядро мембранасына бекінеді, сонымен қатар хроматидтен тұрады. Зиготена синапсисадан, екі гомологтың тығыз конъюгациясынан басталады. Гомологтар конъюгациясынан кейін, олардың ақуызды жіптері жақындап синаптонемальді комплексті түзеді. Конъюгация процессі хромосоманың бірнеше нүктесінен басталады және ұзынан бойлай қосылады. Гомологты хромосоманың конъюгацияға ұшыраған жұптары бивалент деп аталады. Пахитена. Син.комплекс кроссинговерге ұшырайды және алғашқы әртүрлі хроматидтер арасында, бөліктердің алмасуы жүреді. Гендер бір хромосомадан басқа хромосомаға ауысады және жаңа генді комбинация басталады. Диплотенада – синаптонемальді комплекс ыдырайды. Гомологты хромосомалардың биваленттері бір-бірінен ажырайды, бірақта олар кроссинговер жүрген жерде өзара хиазмамен байланысады. Аталық жыныс жасушасында хиазма саны 60-70, ал аналық жасушада -100-ге жуық. Диакинез. Центромера және хиазмамен байланысқан гомологты хромосома хроматидтері бір-бірін итеруі әрі қарай жүреді. Хромосома соңы ядро мембранасынан ажырайды. Диакинез кезеңінде ядро қабықшасы, ядрошық жойылып, бөліну ұршығы · Амитоз (немесе жасушаның тіке бөлінуі) — жасушаның бөлінуі кезінде ұршық түзілмейді. Көп уақытқа дейін митоз және мейозбен бірге жасушаның негізгі бөлінуі болып келді. Қарапайымдылардың, өсімдік және жануарлар жасушаларындағы ядроның бөлінунде кездеседі. · Жасуша амитоздан кейін, қалыпты митоздық циклге өтуге қабылетсіз. · Қазіргі уақытта, амитоз микроскопиялық препараттарды дұрыс дайындамау немесе патологиялық процесс нәтижесі болып саналады. · Амитоз кезінде ядролық қабықша және ядрошық жойылмайды, ядрода бөліну ұршығы түзілмейді, хромоосмалар қалыпты (деспирализденген) жағдайда болады. · Амитоз ядроның және оның кейбір компоненттерінің бірдей бөлінбеуін қамтамасыз етеді. · Эндомитоз, жасуша ядросындағы хромосома санының екіеселенуі, цитоплазма және ядро ары қарай бөлінбейді. Эндомитоз нәтижесінде полиплоидты жасушалар пайда болады, хромосомалық жинақ 2-і есеге жоғарылайды.
1) Гендік инженерия негіздері Бұл ғылымның мақсаты – іс-тəжірибелер мен теория үшін қажетті генетикалық бағдарламаның жобасын жасап, оны тірі организмге енгізу. Мұндай тəсіл тіршілікті меңгеруде жаңа мүмкіндіктер туғызады. генетикалық және биохимиялық әдістердің көмегімен түраралық кедергілері жоқ, тұқым қуалайтын қасиеттері өзгеше, табиғатта кездеспейтін жаңа гендер алу; молек. биологияның бір саласы. Гендік инженерия әр түрлі организмдер геномының бөлігінен рекомбинатты ДНҚ құрастырумен қатар, ол рекомбинатты молекулаларды басқа ағза геномына енгізіп, жұмыс істеуін (экспрессиясын) қамтамасыз етеді. Гендік инженерияның көздеген мақсаты мен зерттеу əдістеріне байланысты мынандай деңгейлерін ажыратуға болады: 1) молекулалық (геннің құрамды бөліктерін зерттеу); 2) гендік; 3) хромосомдық; 4) жасушалық; 5) ұлпалық; 6) ағзалық (организмдік); 7) популяциялық. Басқа биологиялық ғылымдар сияқты генетикалық инженерияның да даму тарихы бар. Бұл саладағы алғашқы зерттеулердің қатарына Н.П.Дубининнің (1934 ж.) дрозофила шыбындарының хромосом санын өзгерту бағытында жүргізген жұмыстары жатады. Ол рентген сəулелерімен əсер ету жолымен хромосома санын өзгертуге болатындығын анықтады. 1956 жылы Е.Сире рентген сəулесінің көмегімен өңделген жабайы эгилопс өсімдігі жапырағының сары дақ ауруына төзімділігін анықтайтын генін, жұмсақ бидайдың хромосомасына апарып салған. Нəтижесінде жұмсақ бидайдың сары дақ ауруына төзімді формасы алынған. ДНҚ құрамынан жеке генді бөліп алу жұмысы тұңғыш рет 1969 жылы Дж. Беквиттің зертханасында жүргізілді. Ол Е.СоІі бактериясынан лактозалық оперон тобына жататын гендерді бөліп шығарды. . Гендік инженериядағы тұңғыш тәжірибені 1972 ж. американ биохимигі Т. Берг (Нобель сыйл. лауреаты) іске асырды. Ол маймылдың онноген вирусы SV-40-тың толық геномын, бактериофаг — L геномының бір бөлігін және Е. Colі бактериясының галактоза генін біріктіру арқылы рекомбинантты (гибридті) ДНҚ алды. 2) Кроссинговер және генетикалық карта Т. Морган және оның зерттеулері, гомологиялық хромосомалар жұбында гендер үнемі алмасып отыратынын көрсетті.Тиісті гендері бар гомологиялық хромосомалардағы сәйкес учаскелердің алмасу процесі хромосомаларыдың айқасуы немесе кроссинговер деп аталады.
Поиск по сайту: |
 АаВв –жаңғақ пішінді
АаВв –жаңғақ пішінді
 – А_B_ -жаңғақ пішінді,
– А_B_ -жаңғақ пішінді,  - А_вв – раушан пішінді,
- А_вв – раушан пішінді,  -аавв-жапырақ пішінді (9:3:3:1)
-аавв-жапырақ пішінді (9:3:3:1) - С_І_ -ақ,
- С_І_ -ақ,  ,
, 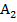 деп белгіленеді.
деп белгіленеді.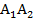 , ал дәннің ақ түсі рецес-і
, ал дәннің ақ түсі рецес-і  ген-мен анықталады. Егер генотипте дом-ы аллельдердің санына тәуелсіз домин-ы аллельдер кездессе, онда дән қызыл түсті болады, ал қалған жағдайда генотиптегі бидай түсі ақ болады.
ген-мен анықталады. Егер генотипте дом-ы аллельдердің санына тәуелсіз домин-ы аллельдер кездессе, онда дән қызыл түсті болады, ал қалған жағдайда генотиптегі бидай түсі ақ болады. – қою қызыл;
– қою қызыл; -қызыл;
-қызыл; - қызғылт;
- қызғылт; - ашық қызыл.
- ашық қызыл. ген-мен бақыланады.
ген-мен бақыланады.